Площади питания плодовых деревьев




Площадь питания
Одним из ведущих факторов в интенсификации плодоводства является плотность насаждений, определяющаяся площадью питания деревьев. Однако для правильного решения проблем в интенсивном плодоводстве важно учитывать не только площадь питания, но и схемы размещения. Установление оптимальных схем размещения деревьев позволяет значительно повысить продуктивность насаждений. В настоящее время в отечественном плодоводстве получили распространение два принципа определения схем размещения. В одном случае уплотнение насаждений достигается путем загущения в ряду при обычных междурядьях, в другом — путем загущения в ряду и сужением междурядий. Значительное увеличение числа высаживаемых на единице площади деревьев обычно сопровождается изменением конструкции сада, введением новых типов формирования и обрезки кроны, применением слаборослых подвоев и сортов. Все больше увеличиваются площади садов с уплотненным размещением деревьев в ряду или с уплощенными вдоль ряда кронами, в результате чего создается непрерывная плодовая стена.
Площади питания плодовых культур зависят от их биологических особенностей, сорта и подвоя, почвенных разностей, рельефа местности, агротехнических условий, влагообеспеченности и направления хозяйства. Плодовые деревья одной и той же породы и сорта, расположенные в разных районах и в неодинаковых условиях, могут сильно отличаться по мощности развития, урожайности и т. п. Кроны деревьев одного и того же сорта яблони в направлении от западных к восточным областям становятся более сжатыми и пирамидальными.
Решающим фактором для интенсивного сада является густота стояния деревьев — количество их на единице площади. По этому показателю различают три типа насаждений. Сады, где на 1 га приходится менее 400 деревьев, относятся к экстенсивным, до 1000 на 1 га — к полуинтенсивным и с количеством деревьев более 1000 на 1 га и формировкой в виде плоского шпинделя или плодовой стены — к интенсивным. Выбор той или иной системы посадки и ведения садоводства определяется социально-экономическими и природно-климатическими условиями. Продуктивность деревьев при увеличении плотности посадки наиболее заметно возрастает лишь до определенного оптимума, соответствующего определенной привойно-подвойной комбинации и конструкции насаждений.
Загущение посадок допустимо только при условии высокого технического уровня формирования крон и обрезки деревьев, обязательного орошения, тщательной обработки почвы и систематического применения удобрений. В противном случае затраты средств не окупаются.
В настоящее время Министерством сельского хозяйства СССР рекомендованы следующие размеры площадей питания (табл. 25, 26).
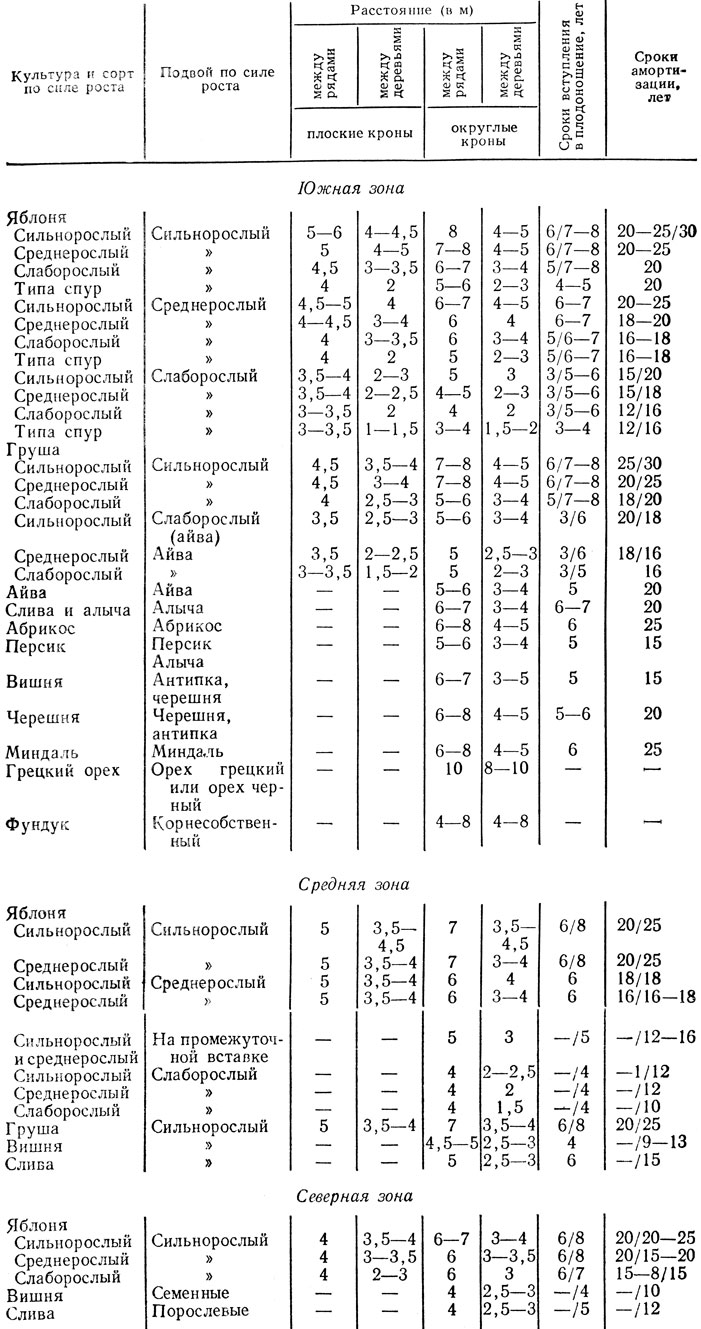
Таблица 25. Площади питания и схемы размещения плодовых деревьев
Примечание. 1. В южной зоне плодоводства насаждения на слаборослых подвоях в районах недостаточного увлажнения исключаются.
2. В северной зоне насаждения на среднерослых и слабо рослых подвоях рекомендуются для производственного испытания.
3. В графе «Сроки вступления в плодоношение» и «Сроки амортизации» в числителе — для плоской формы кроны, в знаменателе — для округлой формы кроны.
Однако надо учитывать, что для одного и того же сорта яблони на одном и том же подвое, но высаженного на разной по содержанию питательных веществ почве, требуется неодинаковая площадь питания. За одно и то же время деревья одного сорта яблони, растущие на бедной песчаной почве, разрастаются гораздо меньше, чем на более богатых суглинистых черноземных почвах.

Таблица 26. Площадь питания и схема размещения ягодных культур
Наилучшее развитие любых плодовых деревьев наблюдается в том случае, если сначала они размещены более густо (обеспечивается взаимная защита деревьев от неблагоприятных условий), а несколько позднее — реже. Поэтому для временного создания более тесного сообщества в рядах, а иногда и в междурядьях высаживают плодовые растения менее долговечных культур, которые завершив полный цикл развития, уступают место более долговечным. Такие временные насаждения называются уплотнителями. Ими обычно являются скороплодные сорта с менее продолжительным сроком жизни. Чаще всего в качестве уплотнителей в разреженном саду высаживают яблоню и грушу. Практика использования уплотнителей в производственных условиях вполне себя оправдала при условии широкой механизированной обработки сада. В интенсивных садах необходимость в уплотнителях отпадает.
Разбивка участка. Разбивку участка под сад проводят в два приема до предпосадочного окультуривания (квартальная разбивка) и после окультуривания (внутриквартальная разбивка). Разбивку начинают со стороны участка, прилегающего к магистральной дороге. Квартальную разбивку сада ведут с помощью геодезических инструментов (теодолит и эккер), пользуются также землемерными лентами и рулеткой, вехами или рейками. Внутриквартальную разбивку сада делают визированием по шнуру или маркированием.
После разбивки квартала на клетки в каждой из них трое рабочих определяют места посадки деревьев визированием. Один из визировщиков становится у предпоследней вешки длинной стороны прямоугольника, установленной при разбивке на клетки, и визирует на предпоследнюю вешку противоположной длинной стороны. Второй становится у второй вешки короткой стороны и визирует на вторую вешку противоположной стороны. Третий ставит вешку на перекрещивании двух линий визировки.
После установки первой вешки первый рабочий остается на месте, второй переходит к соседней вешке, забитой на краевой линии. Затем устанавливают последовательно все вешки первого ряда (третий, четвертый и т. д.). После этого устанавливают вешки во втором и других рядах — по всему участку или кварталу. В конце работы снова проверяют, правильно ли забиты вешки, указывающие местоположение отдельных деревьев, просматривая ряды как под прямым углом, так и по диагонали.
При разбивке по шнуру посадочные колышки расставляют по меткам на шнуре. Вместо посадочных колышков для метки лучше использовать известь, суперфосфат или мел. Разбивка по шнуру нуждается в контрольном визировании, так как возможно смещение шнура из-за разной степени его натягивания.
Маркирование — самый современный способ разбивки больших площадей. Для этого используют широкозахватный культиватор КРН-4,2 или КРН-5,6. На нужных расстояниях по ширине междурядий или расстоянию в ряду устанавливают две лапы — бороздоделатели, а все остальные рабочие органы снимают. Таким маркером сначала размечают расстояния между деревьями в ряду, а затем маркируют и междурядья. Где пересекаются борозды (места посадки деревьев), забивают колышки и визированием проверяют правильность маркирования. В разбивке участка принимают участие тракторист и два рабочих — на провешивании линий.
Для траншейного способа посадки плодовых деревьев разбивку проводят размеченным шнуром. Чтобы в дальнейшем можно было выкапывать траншеи тракторными плугами, разбивочные колья ставят не на месте посадки дерева, а сбоку на расстоянии 1 м от того места, где будет находиться посаженное дерево. Благодаря этому посадочные колья не будут мешать прохождению машин по рядам, что необходимо как для внесения удобрений, так и для выкопки траншеи. При этом посадочные колья надо смещать так, чтобы в одном междурядье они находились один от другого ближе, а в другом дальше. Например, при посадке по схеме 8×4 м в одном междурядье они должны находиться на расстоянии 10 м один от другого, а в другом, соседнем, 6 м. Это обеспечивает челночное движение трактора и других машин по рядам в двух направлениях. Такая разбивка участка называется смещенной.
Для посадки в шахматном порядке сад разбивают специальным треугольником. Он состоит из трех брусков, длина каждого из которых равна принятому расстоянию между деревьями и между рядами. При разбивке сада сначала определяют направление первого ряда и по нему протягивают шнур. Затем треугольник перемещают по шнуру, начиная от места, предназначенного для посадки первого дерева. Каждым положением треугольника определяют места для посадки двух деревьев первого ряда и одного дерева второго ряда. В этих местах вбивают колья. Закончив разбивку первых двух рядов, треугольник переносят к двум первым деревьям второго ряда. Вершина треугольника при этом укажет место для второго дерева третьего ряда. Продолжая таким образом разбивку дальше, обозначают места для деревьев всех рядов участка.
Контурная разбивка очень сложна. Начинают ее с провешивания прямой исходной контрольной линии вдоль склона сверху вниз в наиболее крутой его части. По этой линии забивают колья на расстояниях, соответствующих междурядьям будущего сада. Проделав эту работу по всему склону, приступают к разметке в рядах. Для этого пользуются специальным приспособлением, называемым трассировщиком. Можно пользоваться и нивелиром, но в этом случае затрачивается больше труда. Трассировщик представляет собой рейку длиной, равной расстоянию или половине расстояния между деревьями в рядах. На концах рейки прикреплены две ножки различной длины. Длина ножки зависит от уклона, который желателен для полива сада. Если расстояние между деревьями 6 м и необходим уклон 0,005, то вторая ножка должна быть длиннее первой на 3 см, при уклоне 0,01 — на 6 см. Сверху на рейке около длинной ножки устанавливается уровень.
Начинают разбивку с верхней части склона от первого колышка контрольной линии, проходящей вдоль всего склона, где будет расположен сад. Для этого короткую ножку трассировщика ставят у верхнего первого колышка контрольной линии. Длинную ножку ставят поперек склона так, чтобы рейка заняла горизонтальное положение (определяют уровнем). После этого у длинной ножки забивают колышек, приставляют к нему короткую ножку трассировщика и намечают места для следующих колышков (третьего, четвертого и т. д.) до конца трассы или горизонтального ряда.
Наметив первый ряд, переходят к нижележащему ряду, начав опять от колышка на контрольной линии. Таким путем разбивают все ряды, намеченные на контрольной линии. Если длина рейки трассировщика равна расстоянию между деревьями в ряду, то забитые колышки будут указывать местонахождение плодовых деревьев в ряду. Если длина рейки равна половине расстояния между деревьями, то каждый нечетный колышек явится местом посадки дерева.
Источник статьи: http://agrolib.ru/books/item/f00/s00/z0000040/st067.shtml
Методы определения площадей питания деревьев
В настоящее время лесотаксационная наука располагает достаточно большим количеством методов определения площади питания деревьев. Некоторые ученые (Assmann,1961; Изюмский,1968; Mitscherlich,1970) считают, что за этот показатель дерева в насаждении целесообразно принимать проекцию его кроны. Однако в этом случае между деревьями остаются неучтенные промежутки. По данным многих исследователей площадь их может составлять более 20% (Смертин, 1973; Юодвалькис, 1981; и др.). Они, несомненно, используются корневыми системами, т.к. площади проекций их намного превышают площади проекций крон деревьев. А.К.Поляков (1970) площадь питания деревьев определял как площадь круга с радиусом, равным среднему расстоянию между соседними деревьями, деленному пропорционально развитию их крон .
При определении площади питания деревьев следует руководствоваться следующими предпосылками:
1.Вся площадь насаждения, за исключением больших прогалин и «окон», полностью используется произрастающими на ней деревьями.
2.Каждое дерево в насаждении в процессе конкурентных взаимоотношений с «соседями» использует для своего роста определенную площадь, в соответствии со своими размерами и особенностями пространственного окружения.
3.Площадь питания в известной степени является условной, границы ее в насаждении могут быть выражены не ясно вследствие возможного взаимного перекрытия корневых систем деревьев. В данном случае допускается, что проникновение корней одних деревьев в сферу питания других взаимно компенсируется.
Методы определения площадей питания деревьев и использования их в различных сферах теории и практики лесного хозяйства большое развитие получили в работах зарубежных исследователей. Большинство предложенных методов можно объединить в 4 группы: измерения расстояний между деревьями, построения полигонов (многоугольников) роста, выборочных проб и угловых проб.
Методы первой группы предполагают определение площадей питания деревьев в насаждении только с учетом расстояния между ними. Развитие деревьев и их размеры во внимание не принимаются. Установлено, что среднее расстояние между деревьями находится между значениями расстояний, измеренных от конкретного дерева к третьему и четвертому «соседям» (Prodan, 1965). В этой связи Хаусбург (Hausburg, 1968) разработал уравнение, позволяющие определить число стволов на 1 га на основе среднего расстояния к третьему ближайшему дереву:
N= 10910 L -2.0068 . (1)
Для определение средней площади питания в насаждении предлагается формула:
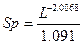 . (2)
. (2)
При определении площади питания конкретного дерева необходимо в последнюю формулу вместо среднего расстояния подставить дистанцию от этого дерева до третьего ближайшего «соседа» (рис. 1 а) .
Построение полигонов роста производиться в двух вариантах. Первый вариант предложен Брауном (Prodan, 1968). Площадь роста исследуемого дерева ограничивается на плане перпендикулярами восстановленными из средин прямых линий, соединяющих данное дерево с его «соседями» (рис.1 б). Таким образом, при проведении натурных работ необходимо от исследуемых (модельных) деревьев измерить расстояния и углы до всех соседних деревьев. Эту работу можно выполнить и по материалам картирования деревьев на пробных площадях.
Второй вариант был разработан Штером (Stоеhr, 1963) и получил дальнейшее развитие и обоснование в работах других исследователей (Prodan, 1968). При определении площади питания искомое дерево соединяется с его ближайшими «соседями» прямыми линиями (рис.1 в ). Каждая линия затем делится на два отрезка, пропорциональных по длине диаметрам деревьев, соединяемых линией

 , (3)
, (3)
где l — определяемое расстояние от исследуемого дерева; li — расстояние от исследуемого дерева до соседнего; d0 — диаметр исследуемого дерева; di — диаметр соседнего дерева.
Через точки деления проводятся перпендикуляры к этим прямым, при пересечении которых образуется полигон площади роста дерева. В данном случае в отличие от первого варианта при сборе данных необходимо дополнительно измерить диаметры всех соседних деревьев. Вейе (Weihe, 1978) считает, что использование при построении полигона роста вместо диаметров деревьев их приростов по площади сечения, существенно уточняет конечный результат.
При определении площадей питания конкретных деревьев путем построения полигонов могут возникнуть сложности с установлением эффективных «соседей». По данным Тяберы (1980), в сосняках II класса бонитета деревья имеют от 3 до 9 соседей. Наши исследования показали, что в сосняках Среднего Урала их количество колеблется от 3 до 11 (Луганский, Нагимов, 1994). Большинство исследователей считают, что среднее количество соседей равно шести, а минимальное их число для получения закрытого полигона — трем. Эти цифры могут служить только ориентиром. В сомнительных случаях следует охватить большее число деревьев. Очень важным преимуществом данного метода является то, что он позволяет при наличии материалов картирования распределить всю площадь насаждения между деревьями.
Большое развитие при определении площадей питания деревьев получил метод выборочных круговых проб (Prodan, 1968; Ehrlenspiel, 1971; Trommer, 1969; Schоеpfer, 1969; и др.). При этом площадь питания устанавливается косвенным путем через определенную на круговых площадках сумму площадей сечений деревьев ( 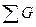 ). Метод используется в трех вариантах: с тремя, шестью и десятью деревьями.
). Метод используется в трех вариантах: с тремя, шестью и десятью деревьями.
Наибольшее распространение получил вариант с шестью деревьями. Поэтому рассмотрим его более подробно. В этом случае для исследуемого дерева находятся шесть ближайших «соседей». Измеряется расстояние до наиболее отдаленного шестого «соседа» (l6). Оно представляет собой радиус круга (выборочной пробы), в центре которого находится исследуемое дерево (рис. 1.4 г). Затем измеряются диаметры всех деревьев, оказавшихся внутри этого круга. По данным выборочной пробы определяется 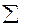 G на 1 га по уравнению
G на 1 га по уравнению
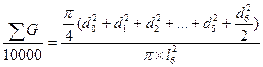 и
и 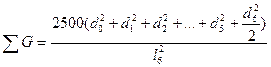 , (4)
, (4)
где d0— диаметр исследуемого дерева; d1…d6 — диаметры соседних деревьев; l6— расстояние до шестого соседа.
Шестое соседнее дерево только наполовину заходит в круг, поэтому при расчете диаметр его учитывается в половинном размере. Площадь питания исследуемого дерева определяется по следующей формуле:
Sp=  , (5)
, (5)
где g — площадь сечения исследуемого дерева.
Варианты выборочных проб с тремя и десятью деревьями не имеют существенных различий от рассмотренного. В этих случаях радиусами выборочных проб являются расстояния от исследуемого дерева, соответственно, до третьего и десятого «соседа».
При работе на угловых пробах, площадь роста деревьев также определяется на основе предварительно установленных значений на 1 га. Штер (Stоеhr, 1968) для этих целей предложил использовать переменный угол визирования, а определять по формуле
=К 2 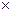 N, (6)
N, (6)
где, К — угол визирования (отчет на шкале, расположенной в 50 см от глаз наблюдателя); N- число учтенных деревьев.
Метод также применяется в нескольких вариантах . Наиболее известны два из них: угловая проба с тремя «соседями» и угловая проба с
Первый вариант предполагает при визировании выбрать такой угол, чтобы охватить три ближайших соседа (рис. 1д). Тогда окажутся учтенными 3.5 дерева, так как только половина третьего соседнего дерева будет входить в площадку. 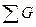 на 1 га определится по формуле
на 1 га определится по формуле
=К 2 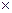 3.5. (7)
3.5. (7)
Для расчета площади питания исследуемого (центрального) дерева используется уже приведенная выше формула. (5).
Второй вариант данного метода предполагает учет десяти соседних деревьев. При этом на 1 га определяется 10 раз. Затем средневзвешенным способом с использованием площадей угловых проб определяется истинное значение на 1 га. Имея этот показатель и площадь сечения дерева, определяется его площадь питания по формуле (5).
Отдельно можно выделить метод определения площади питания деревьев, предложенный Вейе (Weihe, 1976). Он предполагает использование данных перечета деревьев на пробных площадях на основе формулы
Sp= 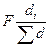 , (8.8)
, (8.8)
где F — площадь насаждения; di— диаметр исследуемого дерева; 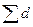 — сумма диаметров всех деревьев.
— сумма диаметров всех деревьев.
При расчетах можно использовать и другие размеры деревьев, например высоту, прирост по площади сечения и т.д.
В насаждении с известными почвенно- климатическими характеристиками площадь питания дерева должна как можно точнее характеризовать условия его роста, слагающиеся из конкурентных взаимоотношений с «соседями». В этом плане предложенные методы, отличающиеся совершенно разными подходами, не могут иметь одинаковые результаты как по затратам труда, так и по точности определения площадей питания деревьев. Так, например, величина площади питания, определенная методом построения полигонов, в значительной степени зависти от характера размещения деревьев. Причем деревья, расположенные в непосредственной близости от исследуемого дерева, но находящиеся за ближайшими «соседями», не участвуют в построении полигонов. Тем не менее нельзя однозначно утверждать, что они не находятся в конкурентных взаимоотношениях с исследуемым деревом. Следует также отметить, что при использовании этого метода не редки абсурдные результаты, когда площади питания деревьев IV и V классов роста оказываются большими, чем деревьев I и II классов. Все остальные методы не учитывают в достаточной мере характер размещения деревьев, а основываются, главным образом, на их удалении от исследуемого дерева. Одни из них при расчете площади питания требует измерения расстояний до всех учитываемых соседних деревьев, другие — только до крайних.
Большинство рассмотренных методов предполагает при определении площади роста использование диаметров или площадей сечений деревьев. В основе их лежит положение о том, что чем больше размеры деревьев, тем больше и приходящиеся им в насаждении площади питания. С этим положением нельзя не согласиться. Однако на основе этих методов нельзя однозначно ответить на вопрос: обусловлены ли размеры деревьев в данный момент определенными для них площадями роста? Это объясняется: во -первых, происходящими с возрастом изменениями пространственного окружения деревьев, которое лежит в основе определения площадей роста; во-вторых, особенностями алгоритма расчета площадей питания большинства методов, предполагающий разделение общей площади насаждения пропорционально размерам деревьев.
Если в конкретном насаждении площадь питания дерева характеризует условия его роста, т.е. поступление света, влаги и элементов питания, то между этим показателем и текущем приростом дерева за какой — то небольшой промежуток времени должна существовать определенная связь. Тогда по тесноте этой связи можно сделать заключение о точности того или иного метода определения площадей питания.
Для изучения этого вопроса нами использовались материалы 4 постоянных пробных площадей, заложенных в 41-48- летних сосняках ягодникового типа леса. На этих объектах были определены площади питания деревьев по всем вариантам первых трех методов. Метод угловых проб по техническим причинам не использовался. Всего этот показатель был определен для 320 деревьев.
В основу исследований положена зависимость текущего прироста деревьев за последние 5 лет по площади сечения на высоте груди от их площади питания. Установлено, что эта зависимость носит криволинейный характер и лучше всего описывается уравнением кривой третьего порядка. Причем теснота связи указанных показателей в значительной степени определяется методом расчета площадей питания.
Метод определения площади питания Корреляционное отношение
1.Расстояний между деревьями 0.25 — 0.35
2.Построения полигонов роста
а) по Брауну 0.16 — 0.37
б) по Штеру 0.58 — 0.82
3.Выборочных круговых проб
а) с тремя деревьями 0.62 — 0.80
б) с шестью деревьями 0.70 — 0.92
в)с десятью деревьями 0.72 — 0.92
4.Метод Вейе 0.60 — 0.84
Как видно из приведенных данных при исследовании зависимости текущего прироста деревьев от их площади питания предпочтение следует отдавать методу выборочных круговых проб с шестью и десятью деревьями. Методы Штера, Вейе и выборочных проб с тремя деревьями обеспечивает несколько меньшую тесноту связи, но вполне приемлемую. Методы расстояний между деревьями и Брауна практически не пригодны для исследований оптимальной густоты древостоев.
Несмотря на то, что выборочные пробы обеспечивают высокую корреляцию прироста деревьев с их площадью питания, они не лишены недостатков. Во — первых, этот метод не обеспечивает полного совпадения абсолютной полноты с истинным значением этого показателя на определенной площади. На исследуемых четырех пробных площадях различия составили от 9 до 15 %. Следовательно, сумма площадей питания всех деревьев не будет равна общей площади насаждения. При некоторых исследованиях такое совпадение крайне желательно. Во — вторых, метод выборочных проб достаточно трудоемкий, особенно его вариант с десятью деревьями.
В этой связи нами предлагается новый метод определения площадей питания. В основе метода лежит положение о том, что при прочих равных условиях деревья в насаждении занимают площади почвенной поверхности в соответствии со своими размерами. При равномерном размещении деревьев этому положению с некоторыми допущениями может соответствовать формула
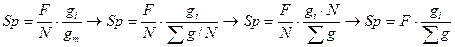 , (9)
, (9)
где gi -площадь сечения исследуемого дерева; gm— средняя площадь сечения древостоя; 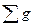 — сумма площадей сечений всех деревьев древостоя; F- общая площадь древостоя; N- количество деревьев в древостое.
— сумма площадей сечений всех деревьев древостоя; F- общая площадь древостоя; N- количество деревьев в древостое.
Данная формула для деревьев одинаковой толщины независимо от их пространственного окружения дает равные площади питания. Это не в полной мере соответствует фактическому положению, особенно в древостоях с резко выраженным неравномерным размещением деревьев. Поэтому для приближения рассчитанной по формуле (9) площади питания к истинной необходимо использовать дополнительный показатель, учитывающий относительное удаление соседних деревьев. Таким показателем может быть высота деревьев. Обобщение литературных (Высоцкий, 1962; Thomasius, Butter, 1984; Вайс, 1995; и др.) и анализ экспериментальных материалов показывает, что варьирование высоты деревьев одинакового диаметра и возраста в значительной степени определяется густотой стояния деревьев (биогрупп ). Таким образом, мы пришли к выводу о необходимости использования при определении площадей питания деревьев показателя g:h:
 , (10)
, (10)
где gi: hi— относительная площадь сечения исследуемого дерева (коэффициент напряженности); 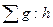 — сумма относительных площадей сечений всех деревьев древостоя.
— сумма относительных площадей сечений всех деревьев древостоя.
Предлагаемая формула позволяет распределить всю площадь древостоя между отдельными деревьями в соответствии с их размерами. Причем площади питания деревьев одинаковой толщины оказываются дифференцированными в зависимости от их высоты. В данном случае высота в определенной степени опосредует влияние пространственного окружения конкретного дерева на величину его площади питания.
Уязвимым меcтом предлагаемого метода может показаться то, что при определении площади питания не учитываются размеры непосредственных соседей. Однако этот недостаток может быть существенным лишь в том случае, если в насаждении существуют строгие зависимости в размещении деревьев разного размера относительно друг друга. Сравнительный анализ полученных материалов показал, что различия в значениях площадей питания деревьев, определенных по формуле (10) и методом выборочных проб не существенны. В то же время предлагаемый метод более прост и устраняет отмеченный недостаток выборочных проб.
Источник статьи: http://helpiks.org/7-58122.html