2. Порядок ржавчинные – Uredinales
Около 4 тыс. видов. Облигатные паразиты, развиваются только на живых частях растения. Мицелий многоклеточный, одноядерный или дикариотический, всегда эндофитный, простирается в межклетниках, внедряясь в клетки гаусториями. Мицелий, как правило, местный, распространяющийся на незначительное расстояние от места проникновения инфекции. Иногда мицелий диффузный, т.е. пронизывающий все органы растения хозяина. Часто мицелий зимующий, многолетний (гаплоидный или дикариотический мицелий). Характерно несколько типов спороношений. В конечном итоге образуются покоящиеся споры или телиоспоры, прорастающие в четырёхклеточную базидию. Телиоспоры могут быть одиночными (одноклеточными, двуклеточными, трехклеточными и многоклеточными), либо развиваться в цепочках.
Ржавчинные грибы могут быть разнохозяинными, когда отдельные типы спороношений развиваются на разных растениях и однохозяинными, когда весь цикл развития гриба проходит на одном виде растения-хозяина.
Цикл развития возбудителя ржавчины злаков – Puccinia graminis (рис. 38). Весной прорастают зимующие на соломе злаков телиоспоры в фрагмобазидию с четырьмя базидиоспорами разных половых знаков (+ и – ). Базидиоспоры активно отбрасываются и попадают в воздушные потоки. Для дальнейшего развития они должны попасть на листья барбариса – промежуточный хозяин. На верхней поверхности листа базидиоспоры (+ и – ) прорастают в гаплоидный мицелий с половым знаком, соответствующим знаку базидиоспоры. Между палисадной тканью и верхним эпидермисом формируются округлые клубочки гиф, которые затем образуют мелкие вместилища с выводным отверстием — пикниды (спермогонии). Спермогонии при созревании принимают форму кувшинчиков, в полости которых от стенок вырастают короткие спороносцы, несущие очень мелкие шаровидные конидии – спермации. Из спермогония наружу выступают короткие гифы — перифизы. К ним прилипают спермации. Между перифизами наружу выступает сахаристая пахучая жидкость с массой погруженных в нее спермациев. Эта жидкость привлекает насекомых, на брюшке и лапках которых спермации переносятся с одной пикниды на другую. Новых заражений спермации не вызывают, но участвуют в половом процессе. Для продолжения цикла развития необходимо объединение двух гаплоидных ядер из + и – спермогониев в дикариотичной гифе.
Объединение спермациев разных половых знаков осуществляется на поверхности листьев барбариса 3 способами.
1) Спермации переносятся с одного спермогония в другой с помощью насекомых.
2) Попавшие на поверхность листа спермации образуют ростковые трубки, которые могут врастать в спермогоний противоположного знака.
3) Содержимое двух ростковых трубок, развившихся из спермациев, может сливаться непосредственно на поверхности листа.
В конечном итоге внутри тканей барбариса развиваются дикариотичные гифы, образующие клубочки внутри тканей листьев, которые затем формируют эцидии в виде чашевидных образований с нижней стороны листьев. В основании эцидия образуется слой базальных клеток, от которого отчленяются в виде цепочек дикариотичные эцидиоспоры гриба. С боков эцидий окружён более толстостенными клетками псевдоперидия.
С изменением ядерной фазы мицелия изменяются паразитические свойства гриба. Эцидиоспоры уже не могут заражать барбарис. Дальнейшее развитие происходит на злаках, куда эцидиоспоры переносятся воздушными потоками. На злаках образуются пустулы в виде порошащих полос или пятен на влагалищах листьев злаков или стеблях. Этот налёт представляет собой следующий тип спороношения — урединии с уредоспорами. Уредоспоры одноклеточные, яйцевидной формы, двуядерные, сидят на клетке-ножке. После разрыва эпидермиса уредоспоры отрываются от своих ножек и способны снова заражать злаки. За лето образуется несколько новых поколений уредоспор (по некоторым данным до семи и более за сезон). На этом же мицелии обычно в конце лета возникают двуядерные телиоспоры с толстой тёмно-бурой оболочкой. Весной каждая клетка телиоспор прорастает четырёхклеточной базидией, куда переходит образовавшееся после слияния двух гаплоидных ядер дикариона диплоидное ядро, делящееся затем редукционно, далее цикл развития повторяется.
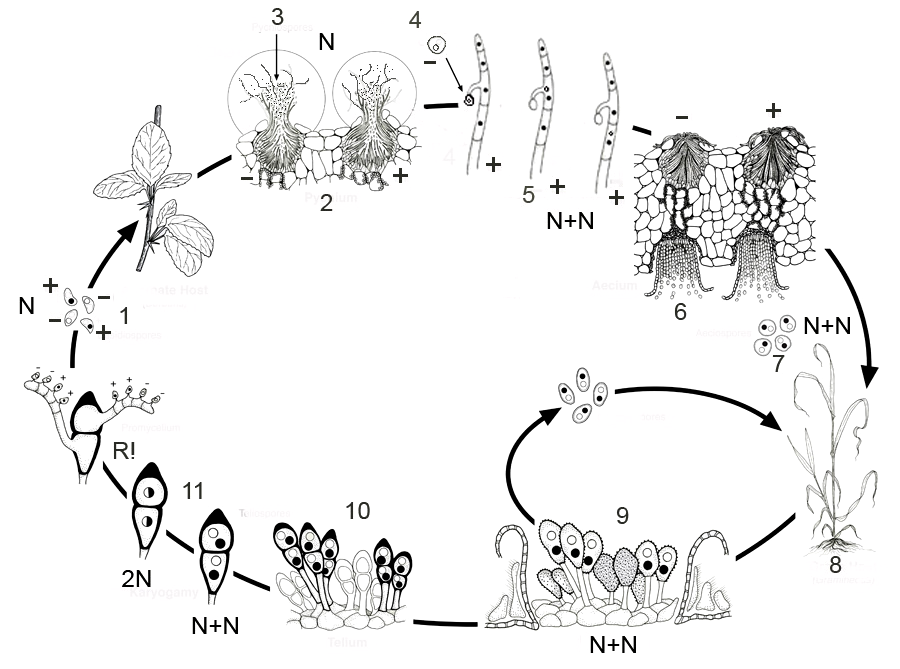
Рис. 38. Цикл развития Puccinia graminis. 1 – базидиоспоры, 2 – спермогонии на листе барбариса, 3 – спермации, 4 – спермация (-), 5 – плазмогамия, 6 – эцидии, 7 – эцидиоспоры, 8 – пораженный злак, 9 – уредоспоры, 10 – телиоспоры, 11 – кариогамия.
Таким образом, в цикле развития ржавчинных грибов имеется пять типов спороношений (табл. 4). Отдельные виды имеют все описанные стадии, другие лишены части спороношений или они у них неизвестны.
Таблица 4. Типы спороношений ржавчинных грибов
Источник
Цикл развития ржавчинных грибов
Как пример разнохозяйственного ржавчинника с полным циклом развития обычно описывают один вид пукцинии (Pиcciпia gramiпis), вызывающий линейную ржавчину злаков. Весной развитие гриба начинается на листьях барбариса, где обнаруживаются небольшие гипертрофированные оранжевые участки ткани, пронизанные межклетным мицелием с гаусториями. Мицелий — этот состоит из одноядерных клеток с гаплоидными ядрами; оранжевый цвет зависит от окрашенных капель масла в его клетках. На пятнах образуются два погруженных в ткань спороношения гриба: на верхней стороне листа.- пикниды, или спермагонии, на нижней стороне – эцидии. Пикниды имеют кувшинообразную форму ; в полость от стенок врастают короткие конидиеносцы, отчленяющие мелкиеодноядерные конидии, называемые пикноспорами или спермациями. Наружу из пикниды высовывается пучок гиф и выделяется пахучая сахаристая жидкость, в которую погружены пикноспоры; вызвать новую инфекцию пиктоспоры не могут. Эцидий закладывается в глубине листа в виде клубка гиф из одноядерных клеток. Затем в базальной части его образуется компактная площадочка двухъядерныx цилиндрических, так называемых базальных клеток, кoтoрые отчленяют базипетальные цепочки двухъядерных, материнских клеток спор, каждая из последних делится на верхнюю эцидиоспору с оранжевым содержимым — и нижнюю, маленькую, позднее разрушающуюся промежуточную клетку. Периферические базальные клетки образуют подобным же образом оболочку эцидия – перидий, -окружающую его с боков; наружные эцидиоспоры превращаются в клетки, образующие крышку перидия, смыкаются с его боковыми стенками. Эцидии отчасти выдаются над нижней поверхностью листа; перидий эцидия на морфологической вершине его, или крыше, разрывается, и лопасти его в виде зубчиков отворачиваются наружу, так что эцидий получает вид вазочки, наполненной эцидиоспорами. Последние разъединяются вследствие разрушения промежуточных клеток, выпадаютJ вниз из эцидия и разносятся воздушными течениями. Они не могут заражать вновь барбарис, а заражают листовые влагалища и стебли злаков. Там они развивают межклеточный мицелий, состоящий из клеток с дикарионами. Через 5-7 дней на нем под эпидермисом растения-хозяина образуется новое спороношение гриба — собрание овальных одноклеточных двухъядерных уредоспор, сидящих каждая на ножке. Эпидермис растения-хозяина раскрывается, и обнажаются уредоспоры, собранные в заметные невооруженным глазом ржавые полоски, несколько похожие на ржавчину на железе. Окраска зависит от оранжевых капель масла в клетках уредоспор. Разносимые воздушными течениями, уредоспоры прорастают и вызывают новое заражение злаков; такие заражения повторяются 5-6 раз в лето, тк что от первичноно заражения злака одной эцидиоспорой потенциально к концу лета можетобразоваться 10 15 — 10 18 уредоспор. К концу лета нa том же мицелии, который давал уредоспоры, образуется новое спороношение – телейтоспоры : они также сидят на ножках и состоят каждая из двухдвухъядерных клеток с толстой темно-бурой оболочкой. На злаках скопления телейтоспор заметны в виде черных продолговатых полосок на влагалищах листьев и на стеблях. Телейтоспоры прорастают только следующей весной, после перезимовки. Перед этим ядра дикарионов сливаются; получившиеся диплоидные ядра делятся редукционно, и из каждой телейтоспоры вырастает четырехлеточная фрагмо6азидия% на каждой клетке фрагмобазидии образуется по одной гаплоидной одноядерной базидиоспоре. Базидиоспоры, разносимые воздушными течениями, заражают листья барбариса, образуя в них гаплоидный мицелий, и цикл развития повторяется. Весь цикл развития Pиcciпia gramiпis и смена ею растений-хозяев были впервые выяснены работами немецкого миколога А. Д е — Б ари в 1864-1865 гг. В течение долгого времени была неясна роль пикноспор, и только В 1927 г. канадский миколог и фитопатолог Крэги выяснил их значение. Его опыты обнаружили, что ржавчинники гетероталличны и что на гаплоидном мицелии (+ или -), развивающемся из заражения одной базидиоспоорй, не могут образоваться эцидиоспоры, содержащие по 2 ядра (дикарионы). Для образования их необходимо наличие еще ржавчинника противоположного полового знака. В природе это осуществляется или при срастании мицелиев противоположных знаков, получившихся на одном листе барбариса вследствие заражения несколькими (хотя бы двумя) разнозначными базидиоспорами, или при переносе насекомыми пикноспор одного знака в пикниды мицелия противоположного знака. Лучшему улавливанию этих пиктоспор способствуют особые длинные извилистые гифы, высовывающиеся из пикнид: к ним прилипают пикноспоры, переносимые насекомыми. Содержимое пикноспоры переходит в эти гифы и далее по ним в мицелий. Ядра пикноспор, очевидно, размножаются делением и через некоторое время образуют с ядрами мицелия, куда они попали, дикарионы, после чего становится возможно образование двухъядерных эцидиооспор. Цитологические подробности этой дикарионтизации еще недостаточно выяснены. Таким образом, в полном цикле развития ржавчинного гриба имеются: 1) гаплоидная фаза, начинающаяся с базидиоспор и представленная далее гаплоидным мицелием с пикнидами и зачатками эцидиев (только зачатками, но без эцидиоспор); 2) дикарионтическая фаза, начинающаяся незадолго до образования эцидиоспор, длительно продолжающаяся затем через большую часть цикла развития с повторным образованием дикарионтического мицелия с уредоспорами и заканчивающаяся в телейтоспорах перед слиянием их ядер; 3) диплоидная фаза в виде телейтоспор с диплоидными ядрами до редукционного деления их.
1. Введение в культуру устойчивых сортов цветочных растений.
2. Обрезка пораженных побегов и стеблей, уборка опавших листьев и других остатков, с последующим сжиганием их или закладкой в компостные кучи.
3. Перекопка или перепашка почвы на глубину пахотного слоя, с оборотом пласта.
4. Опрыскивание 1-процентной бордосской жидкостью в первые дни отрастания цветочных растений, что позволит не допустить заражения, вызываемого базидиоспорами. В дальнейшем против стадии уредоспор следует обрабатывать растения до цветения препаратами серы.
5. При развитии ржавчины на побегах розы, с образованием на них разрывов коры и стадии эцидиоспор, места поражения следует зачищать и смазывать раствором нафгената меди или 3-процентным раствором медного купороса. Хорошие результаты дает смазывание мест поражения двухлетних и многолетних побегов 50-процентным карболинеумом, a также опрыскивание роз перед укрытием на зиму 8—10-процентным карболинеумом или 4—5-процентной бордосской жидкостью.
6. Уничтожение на плантациях сорной растительности. В борьбе с ржавчиной роз не допускать поблизости произрастания шиповника, а в борьбе с ржавчиной астр и пеона — обыкновенной сосны и сосны ригида. При разведении гвоздики вести борьбу с молочаем.
7. Создание вокруг цветочных плантаций защитных полос из древесных пород, не поражаемых ржавчиной, в целях преграждения заноса спор со стороны.
Источник