Маревые ландшафты что это
Рельеф Хехцира еще более ста лет назад охарактеризовал известный натуралист и путешественник Р.К. Маак (1861), назвавший этот горный массив “интересным во всех отношениях”. Он писал: “За исключением некоторых скалистых обнажений, находящихся вблизи устья Чирку, хребет гор, который носит название Хехцыр, имеет склоны отлогие, незаметно восходящие к вершине и покрытые мешанным лесом, между тем как на гребнях этих гор снова появляются зубчатые скалистые обнажения” (1861, с. 45).
Среди всех островных гор среднего течения Амура Хехцир выделяется наибольшей высотой (до 950 м над ур. моря) и тем, что он представляет самостоятельный антиклинорий, т.е. складчатую структуру, изгибом обращенную вверх, с выходом на поверхность более древних слоев в центральной части. Хехцирский антиклинорий сложен сильно дислоцированными верхнепалеозойскими кремнистыми сланцами и мезозойскими глинистыми сланцами, песчаниками и конгломератами. Отмечены здесь и кварцевые диориты (Никольская, 1958).
Рельеф заповедника по преимуществу горный, однако здесь имеются и равнинные участки – плоские, слаборасчлененные поверхности древнеозерных террас. Они лежат на абсолютных высотах от 35 до 100 м, в среднем 40–50 м над ур. моря. Для равнинных участков характерны длительносезонная мерзлота и переувлажненность; основной тип ландшафта – маревый. Предгорья Хехцира – холмисто-увалистые. Средние высоты – 80–150 м, отдельные холмы достигают 200–250 м н.у.м. Расчлененность рельефа средняя, густота речной сети – 0,6 км/км 2 . Ширина долин – от нескольких десятков до нескольких сот метров; их плоские днища переувлажнены. На большей части заповедника преобладает средневысотный горный рельеф с мягкими, сглаженными очертаниями. От осевого хребта, вытянутого в широтном направлении, на север и юг отходят отроги высотой 300–500 м над ур. моря. Густота речной сети здесь достигает 0,8 км/км 2 . Гребни водоразделов местами узки и каменисты, встречаются отвесные скальные уступы. Эти формы рельефа связаны обычно с выходами кварцевых порфиров.
Все реки заповедника принадлежат бассейнам Уссури и Амура. Самая крупная река – Чирка имеет в длину 82 км и течет, чрезвычайно сильно петляя, вдоль подножия Хехцира с востока на запад. Длина большинства рек, стекающих с хребта (Одыр, Цыпа, Белая, Быковая, Пилка), составляет всего 8–17 км. Реки мелководные, в верхнем течении имеют типично горный характер, в нижнем – равнинный, с низкими, часто заболоченными берегами и многочисленными меандрами. Есть несколько старичных озер – Нимфейное, Сюрприз, Бразениевое. Весенние паводки не выражены, наводнения обычны летом, в период выпадания муссонных дождей.
По климатическому районированию Дальнего Востока Г. Н. Витвицкого (1969), территория заповедника относится к Амуро-Уссурийскому климатическому району, который входит в Тихоокеанскую муссонную область умеренной зоны. Климат в целом характеризуется как летне-теплый (сумма температур за вегетационный период 2500°С) и влажный (индекс сухости 0,45–1,00) с умеренно суровой малоснежной зимой. Климатические показатели в разных частях заповедника в зависимости от высоты над уровнем моря, экспозиции, крутизны и протяженности горных склонов, ветрового режима и температурных инверсий различаются между собой. Наиболее благоприятны климатические условия в предгорной местности и в среднем горном поясе до высоты 400–500 м н.у.м. Здесь средняя годовая температура 1,4°, продолжительность безморозного периода –150-160 дней, вегетационного – 140–-150. Средняя температура января – 1-18°, июля 19-20°. Сумма положительных температур за вегетацию 2400–2500°. Общая годовая сумма осадков – 600-700 мм.
В верхнем горном поясе (выше 600 м над ур. моря), где распространены пихтово-еловые и каменно-березовые леса, климат более суров и контрастен. Средняя годовая температура воздуха –3,2°, средняя температура января –20–23°, июля 14–16°, продолжительность безморозного периода – 110–120 дней, вегетационного –90–100. Сумма положительных температур за вегетацию 1300–1400°. Годовая сумма осадков – 900–1000 мм. Так же суровы местные климатические условия и на равнинных участках, где безморозный период почти на месяц короче, чем в предгорьях и в среднем поясе гор. На участках маревого ландшафта под заболоченными лиственничниками и сырыми закочкаренными вейниково-осоковыми лугами имеется длительносезонная мерзлота.
Под низинными и переходными болотами с лиственницей развиты торфяно-глеевые почвы. Мощность торфяного горизонта в них достигает 0,5 м и более. На плоских, пониженных участках долин рек отмечены задернованные иловато-глеевые почвы. Эти почвы сильно переувлажнены и имеют грубозадернованный верхний горизонт мощностью до 5–7 см (Васильев, 1979). Ниже залегает темно-серый перегнойный горизонт мощностью 15–20 см, переходящий в глинистый глеевый. Подстилаются эти почвы хорошо окатанной галькой. Грунтовые воды отмечаются на глубине 40 – 50 см. В период муссонных дождей низинные участки иногда на продолжительный срок покрываются слоем воды. Задернованные иловато-глеевые почвы имеют слабокислую реакцию среды, в нижних горизонтах даже близкую к нейтральной, содержат большое количество гумуса и богаты азотом и подвижными соединениями фосфора. Потенциальное плодородие этих почв высокое. К подобным местообитаниям приурочены заболоченные лесные сообщества из ольхи пушистой, лиственницы Гмелина и ясеня маньчжурского.
Под дубовыми и березово-осиновыми лесами на релках развиты бурые, в различной степени оглеенные почвы. Для этих почв характерен сильно задернованный гумусовый горизонт мощностью 10–15 см, который сменяется несколько осветленным элювиальным горизонтом мощностью 30–40 см. Под кедрово-широколиственными и широколиственными лесами на горных склонах и в предгорьях развиты бурые горно-лесные почвы. Они наиболее характерны для местообитаний с хорошим дренажем. Имеющийся на поверхности этих почв свежий опад почти полностью минерализуется в течение вегетационного периода.
К пихтово-еловым лесам верхнего пояса гор приурочены буро-таежные иллювиально-гумусовые почвы. Толщина слоя полуразложившейся лесной подстилки составляет 3–5 см. Гумусовый горизонт рыхлый и содержит большое количество свежих и полуразложившихся растительных остатков, его мощность – 10–12 см. Ниже залегает рыхлый коричневатый иллювиальный горизонт. У этих почв сильнокислая реакция среды и высокая емкость обмена. В гумусовом горизонте происходит накопление поглощенных оснований кальция, магния, а также натрия, титана, фосфора и марганца. В иллювиальном горизонте отмечено высокое содержание подвижных форм железа и алюминия. Для буро-таежных иллювиально-гумусовых почв характерно грибное разложение растительных остатков. Довольно часто наблюдается большое количество грибов в виде грибницы и нитей мицелия по всему почвенному профилю.
В верхнем поясе гор под каменноберезниками развиты лугово-лесные почвы. Характерная их особенность – грубогумусовый горизонт, достигающий мощности 20–25 см и содержащий большое количество средне- и слаборазложившихся растительных остатков. Реакция среды слабокислая, близкая к нейтральной. В нижней части почвенного профиля часто встречаются темные гумусированные линзы, прослойки и гумусовые затеки по морозобойным трещинам, достигающие глубины 40–50 см.

© Н.Г. Васильев, Е.Н. Матюшкин, Ю.В. Купцов. Большехехцирский заповедник. // Заповедники СССР. Заповедники Дальнего Востока. — М, Мысль, 1985.
Источник статьи: http://oopt.info/index.php?oopt=1202
Маревые ландшафты что это




Семейство маревые (Chenopodiaceae) (В. И. Грубов)
Маревые ассоциируются в сознании большинства людей с злостными садово-огородными сорняками и рудеральными растениями. II это соответствует действительности: виды мари (Chenopodium) и лебеды (Atriplex) — распрост-раненнейшие сорняки-космополиты. Но вместе с тем этому семейству человечество обязано корнеплодом первостепенной важности — свеклой и прежде всего сахарной свеклой, которая ныне дает более трети всего производимого в мире пищевого сахара, а также повседневным овощем — столовой свеклой. Кроме того, это семейство содержит ряд других менее известных, но ценных полезных растений, о чем будет речь далее.
Внешне маревые — невзрачные растения: среди них почти нет декоративных видов, введенных в культуру. Но при произрастании в массе, на большом пространстве некоторые из них могут создавать красочный покров. Так, в пустыне Гоби поташник стройный (Kalidium gracile, табл. 58), образующий почти чистые сообщества на глинистых такырах, изумрудно-зеленый в летний период, осенью окрашивается в яркий янтарно-желтый или малиново-красный цвет, создавая на белесом фоне пустыни издалека бросающиеся в глаза обширные цветные пятна. В пустынях некоторые виды маревых создают осенние аспекты за счет ярко окрашенных и сверкающих на солнце полупрозрачных крыльев плодов, например солянка мясистая (Salsola crassa), солянка казахов (S. ka-sakorum) и ежовник безлистный (Anabasis aphylla, табл. 58).
Среди маревых есть однолетние и многолетние травы, полукустарнички и кустарники и даже небольшие деревья (саксаул — Haloxylon, табл. 59). Стебли обычно прямостоячие, реже простертые. Только у кавказского лесного монотипного рода габлиция (Hablitzia) да еще у двух кустарниковых видов из близкородственных между собой родов: рагодия (Rhagodia) из Австралии и хольмбергия (Holmbergia) из Южной Америки — стебель лиановидный, лазящий. У кавказского вида габлиции тамусовидной (Hablitzia tamnoides) он травянистый, ежегодно отмирающий, достигает в длину 2 м (рис. 203).
Для многих маревых характерна членистость стебля и ветвей, хорошо выраженная, например, у солероса (Salicornia) и саксаула. При этом нередко наблюдается редукция листьев, от которых остается лишь очень короткое влагалищное кольцо со следами листовых пластинок в виде бугорков или шипиков. Некоторым маревым свойственны колючие окончания ветвей, которые наблюдаются как у кустарниковых видов (например, в родах ноэа — Noaea, табл. 59, рагодия), так и у травянистых (акроглохип — Acroglochin, телоксис — Teloxis), или же колючие окончания листьев (у многих видов солянки, ежовника и других родов).
Среди пустынных полукустарничковых маревых, в том числе и у членистостебельных, часто наблюдается подушковидная форма роста. Плотные подушки образуют в пустынях Казахстана тасбиюргун (Nanophyton erinaceum) и саксаульник длипноприцветниковый (Arthrophytnm longibracteatum), в песках Каракумов и Кызылкума — саксаульник Лемана (A. lehmannianum), в каменистых пустынях Сахары — фредолия аретиевидная (Fredolia aretioides). Плоские лепешковидные или ковриковидные куртины образуют биюргун (Anabasis salsa), камфоросма Лессинга (Camphorosma lessingii), саксаульник бетпакдалинский (Arthrophytum betpakdalense).
Листья маревых без прилистников, очень разнообразные по форме — от шиловидных и линейно-цилиндрических жестких до широких, овальных, мясистых, цельнокрайних, зубчатых, иногда неглубоко лопастных. Листорасположение как очередное (преобладающее), так и супротивное (особенно у членистостебельных маревых).

Рис. 203. Габлициятамусовидная (Hablitziatamnoides): 1 — часть цветущего побега; 2 — плоды с околоцветником (один открывшийся с отпавшей крышечкой и выпавшим семенем, другой открывающийся); 3 — семя
Корень большей частью стержневой, разветвленный, но у некоторых родов имеется тенденция к его разрастанию, утолщению и превращению в запасающий орган. Эта тенденция наиболее выражена у свеклы, что и было использовано человеком для выведения ее культурных сортов. Веретеновидные и клубневидные корни встречаются у видов бассии (Bassia), камфоросмы, мари, сведи (Suaeda). У лианы габлиции тамусовидной корень неправильно-клубневидный, бугорчатый.
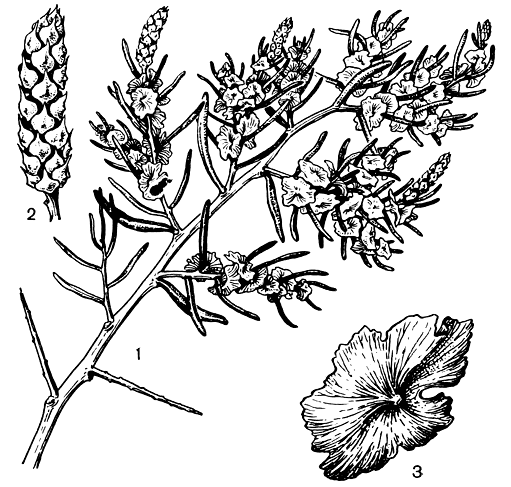
Рис. 204. Саркобатус червелистный (Sarcobatus vermiculatus): 1 — ветвь с плодами и мужскими колосками; 2 — мужской колосок; 3 — плод
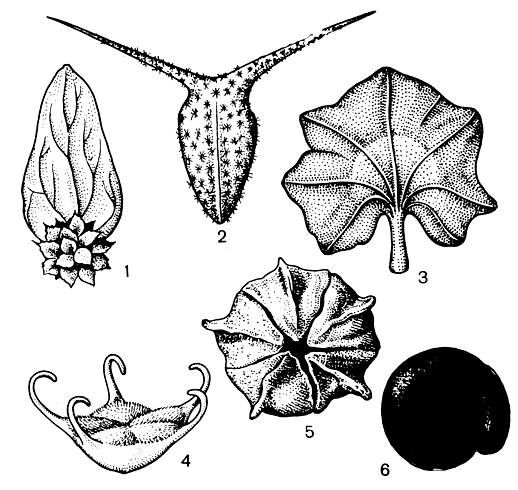
Рис. 205. Плоды и семена маревых: 1 — шпинат огородный (Spinacia oleracea) — соплодие; 2 — рогач песчаный (Ceratocarpus arenarius) — плод; 3 — лебеда вееро-плодная (Atriplex flabellum) — плод; 4 — бассия иссополистная (Bassia hyssopifolia) — плод; 5 — 6 — марь белая (Chenopodium album) — плод (5) и семя (6)
Тычинок обычно столько же, сколько и чашелистиков, и они всегда противостоят им. Реже тычинки в меньшем числе и даже одна, как у рогача и некоторых видов верблюдки (Corispermum), петросимонии (Petrosimonia). Нити тычинок свободные или сросшиеся при основании, образующие подпестичный диск. Пыльники нередко бывают ярко окрашенными (желтыми, красными, розовыми) и довольно крупными. Когда они при полном цветении сильно выставляются из околоцветника, то создают иллюзию окрашенного цветка. У некоторых маревых они снабжены на верхушке придатками различной формы, иногда пузыревидно вздутыми и более крупными, чем пыльники, ярко окрашенными (розовыми, белыми, желтыми), создающими такой же эффект. Цветки маревых могут быть и обоеполыми и раздельнополыми даже на одном и том же растенииу и в таком случае верхние цветки в соцветии мужские, а нижние женские, но растения могут быть и двудомными и полигамными. Своеобразно устроено мужское соцветие у эндемичного североамериканского рода саркобатус (Sarcobatus, рис. 204), стоящего особняком в семействе маревых: тычинки с очень короткими нитями и без околоцветника, без особого порядка распределены под грибовидными щитками, образующими довольно плотное верхушечное колосовидное соцветие. Женские цветки одиночные, пазушные, со спайнолистным околоцветником, почти полностью сросшимся с завязью и при плодах развивающим широкое складчатое кольцевое крыло.
Гинецей обычно из 2, реже 3 — 4 или даже 5 плодолистиков, со свободными или более или менее сросшимися столбиками. Завязь верхняя или редко полунижняя (свекла) с одним базальным семязачатком. Плод нераскрываю-щийся, с пленчатым околоплодником, большей частью окруженный остающейся чашечкой и вместе с ней опадающий. Редко околоплодник мясистый, сочный и тогда плод ягодовидный, как у австралийской рагодии, или, напротив, твердеющий и тогда плод открывающийся крышечкой, как у свеклы и габлиции. У солянки, саксаула, ежовника и других, близких к ним родов, околоцветник ко времени созревания плода развивает по периметру широкие, заходящие друг за друга полупрозрачные крыловидные придатки, то бесцветные, то окрашенные в различные яркие цвета — золотисто-желтые, лимонно-желтые, оранжевые, малиново-красные, дымчатые и т. п. И в этот период названные растения очень красивы, при массовом развитии они создают яркие аспекты. Таким образом невзрачность цветков у многих маревых компенсируется красочностью плодов. У некоторых родов (кохии — Кochia, кириловии — Kirilowia, лондезии — Londesia) околоцветник покрыт длинными спутанными белыми волосками и плоды тогда выглядят шерстистыми клубочками в пазухах листьев. У лебеды, рогача, шпината в образовании плода участвуют прицветнички, часто срастающиеся частично или полностью и сильно разрастающиеся, как, например, у лебеды монетоплодной (Atriplex moneta) или лебеды веероплодной (A. flabellum, рис. 205). Иногда благодаря срастанию между собой околоцветников в соцветии, как у свеклы, или прицветничков, как у шпината, образуются соплодия (рис. 205).
Для систематики маревых большое значение имеет форма покрывающих их волосков, поскольку она характерна для целых таксономических групп внутри семейства. Так, мари, лебеде и близким к ним родам присущи воздухоносные пузыревидные волоски (отчего листья и ветви этих растений имеют мучнистый или серебристый покров); камфоросме, солянке и близким родам — членистые и часто зазубренные длинные волоски; верблюдке (Corispermum) и родственным родам — ветвистые волоски; группе терескеновых (терескен, рогач, аксирис и др.) — звездчатые волоски, часто с длинным лучом в центре; родам петросимония и офайстон (Ofaiston) — двуконечные (мальпигиевые) волоски; группе солянок, близких к солянке почечконосной (Salsola gemmascens), — двуконечные прозрачные чешуйки.
В семействе насчитывается немногим более 100 родов и около 1500 видов. Семейство маревых довольно четко разделяется на два подсемейства: маревых (Chenopodioideae), у всех видов которого семена имеют кольцевидно согнутый зародыш и развитый эндосперм, и солянковых (Salsoloideae), у которых зародыш закручен в спираль и эндосперм не развит. Эти два подсемейства далее подразделяются на несколько групп, в отношении таксономического ранга и границ которых мнения различных систематиков расходятся. Более или менее признанными можно считать следующие 8 триб.
К подсемейству маревых относятся 5 триб: хруплявниковые (Polycnemeae), свекловые (Beteae), маревые (Chenopoclieae), верблюдковые (Corispermeae) и солеросовые (Salicornieae); подсемейство солянковых включает 3 трибы: саркобатовые (Sarcobateae), сведовые (Suaedeae) и солянковые (Salsoleae).
Наиболее крупными родами являются марь (около 250 видов), лебеда (около 220), солянка (свыше 200) и сведа (до 100); 42 рода монотипны, т. е. содержат по одному виду. Большинство родов (42) и видов (свыше 850) относятся к трибе маревых. Далее следуют солянковые (около 30 родов и 350 видов), сведовые (5 родов и около 100 видов) и солеросовые (12 родов и более 60 видов). Остальные 4 трибы насчитывают от 1 до 5 родов каждая и около 30 видов все вместе.
Маревые распространены по всему земному шару от полярной зоны до тропиков. Ряд родов — марь, лебеда, кохия, солерос, артрокнемум (Arthrocnemum) и сведа — представлен на всех континентах — в Евразии, Северной и Южной Америке, Африке и Австралии. Это говорит о том, что семейство маревых сформировалось в основном еще тогда, когда между континентами сохранялась непосредственная связь, следовательно, не позднее начала мелового периода. Однако достоверные следы маревых известны лишь с третичного периода.

Рис. 206. Маревые. Кумарчик колючий (Agriophyllum pungens): 1 — часть растения; 2 — плод. Солерос европейский (Salicornia europaea): 3 — общий вид растения; 4 — цветок
Существуют 7 главных областей распространения, многообразия и эндемизма маревых:
1) низменные пустынные территории Австралии, особенно ее южной части. Это область наибольшего богатства и разнообразия маревых. Эндемизм здесь достигает 92% (из 130 зарегистрированных видов 120 — эндемики). 20 родов встречаются только в Австралийской области;
2) пустынные и засоленные прибрежные территории Средиземного моря, включая пустыни Северной Африки. Здесь можно насчитать 15 эндемичных родов. Род свекла свойствен этой области;
3) пустыни и степи Передней и Средней Азии имеют 16 «своих» родов;
4) пустыни Центральной Азии. Здесь маревые играют ведущую роль в растительном покрове, многие из них определяют ландшафты пустынных территорий и около 10 родов специфичны для этой области.
Три последние перечисленные области непосредственно прилегают друг к другу, тесно связаны и имеют много общих родов;
5) пустыни и прерии Северной Америки. Этой области присущ настолько своеобразный род саркобатус (2 вида), что его выделяют в особую трибу. Кроме того, еще 8 родов эндемичны;
6) пустыни и пампа Южной Америки. Отсюда известны 3 эндемичных и 2 общеамериканских рода и множество эндемичных видов мари и лебеды;
7) Южная и Юго-Западная Африка. Здесь в пустынях Намиб и Карру распространен особый род из лебедовых — экзомис (Exomis) — и целый ряд эндемичных видов солянки, а так же род хенолея (Chenolea), присущий и пустынным районам Южной Африки.
Как видно из перечисления основных районов распространения маревых, это обитатели аридных и сильно засоленных территорий. Действительно, за исключением сорных и рудеральных растений, это преимущественно ярко выраженные ксерофиты и классические галофиты, живущие часто в условиях крайней сухости и чрезвычайного засоления почвы, которых уже не выносят никакие другие растения. Например, илъиния Регеля (Iljinia regelii) из солянковых является единственным обитателем выжженных солнцем и сильно загипсованных каменисто-щебнистых делювиальных шлейфов гор в Западной Гоби, где осадков выпадает до 40 мм, и то не каждый год. Там же, в Гоби, встречается полукустарничек из солянковых симпегма Регеля (Sympegma regelii), который растет на совершенно голых, черных от пустынного загара скалах сопок и предгорий.
Соляноколосник каспийский (Halostachys caspica) и сарсазан (Halocnemnm strobilaceum) из солеросовых растут на пухлых и мокрых солончаках, где соль выступает в кристаллах большими белыми пятнами и ничто другое уже расти не может, а солерос европейский (Salicornia europaea, рис. 206) часто растет прямо в концентрированном соленом растворе по закраинам соленых озер. Как победители в борьбе за существование в экстремальных условиях сухости и засоленности почвы, сильного перегрева, резких суточных колебаний температуры, интенсивной инсоляции, иссушающих сильных ветров, многие маревые являются эдификаторами в пустынях.
На так называемых «черных землях» юго-востока европейской части нашей страны и в пустынях Казахстана широко распространены солянково-полынные пустыни, где наравне с по дынями выступают или биюргун, или итсегек (Anabasis aphylla, табл. 58), или тасбиюргун, или камфоросма монпелъенская (Camphorosma monspeliacum). Аналогичные солянково-полынные пустыни развиты в Северной Америке на юге США, где партнерами полыни трехзубчатой выступают или кохия одетая (Kochia vestita), или лебеда скученно листная (Atriplex confertifolia), или саркобатус червелистный (Sarcobatus vermiculatus), в зависимости от формы и степени засоленности и влажности почвы. В Северной Африке подобные пустыни образуют полынь белотравная и ежовник членистый (A. articulata), а также лебеда.
В пустынях маревые часто завоевывают уже полное господство, образуя чистые сообщества или доминируя в смешанных. Вспомним, например, знаменитые саксауловые леса в песчаных пустынях нашей Средней Азии и Казахстана, Передней и Центральной Азии, а также низкорослые разреженные саксаульники в щебнистых пустынях Гоби (карта 17).
Широкое распространение имеют пустыни с жестко- и колючекустарничковыми маревыми. В Северной Африке они слагаются хенолеоидесом арабским (Chenoleoides arabica), солянкой четырехугольной^ alsola tetragona), лебедой настоящей (Atriplex halimus) с участием крупного однолетника — бассии колючковой (Bassia muricata). В Бетпак-Дала и пустынях Восточного Казахстана это боялычники, где господствуют солянка деревцевидная, или боялыч (Salsola arbuscula), либо солянка боялычевидная (S. arbusculiformis). В Центральной Азии такие же сообщества на больших площадях образуют солянка лиственницелистная (S. laricifolia) и тот же боялыч. В пустынях Австралии (на северо-востоке и юге) подобные сообщества слагают лебеда пузырчатая (Atriplex vesicaria) и виды кохии. В Северной Африке, в алжирской щебнистой пустыне, где к неистовому солнцу и страшной сухости присоединяется постоянный сильный ветер, полукустарничек фредолия аретиевидная, близкий к роду ежовник, образует низкие плотные подушки диаметром до 120 см, которые на огромных пространствах создают своеобразный мрачный ландшафт. Аналогичные подушечники на юге Африки, в суровых пустынях Карру и Намиб, создают виды солянки.
Маревые — основные обитатели мокрых и пухлых солончаков по морским побережьям и по берегам временных и постоянных соленых внутриконтинентальных озер. Это безлистные суккуленты из трибы солеросовых и многочисленные также суккулентные сведы.
Широчайше распространен сарсазан, образующий огромные, плоские, ярко-зеленые подушки-куртины на влажных и пухлых солончаках Средиземноморья от Марокко и Испании до Передней и Средней Азии, в Центральной Азии от Казахстана до подножия Наныпаня. Выше уже упоминался соляно-колосник, очень характерный для пухлых и мокрых солончаков равнин Передней и Средней Азии и Казахстана. Он растет довольно крупными кустами, большими зарослями, чистыми или с участием тамариска. В Северной Америке подобные заросли на влажных и мокрых солончаках создают алленролъфея западная (Anenerolfea occidentalis), а в Австралии — артрокнемум деревце видный (Arthrocnemum arbuscula) и артрокнемум сареазановидный (A. halocnemoides), пахикорния тонкая (Pachycornia tenuis).
На влажных морских побережьях Евразии широко распространены сообщества артрокнемума сизого (Arthrocnemum glaucum), солероса кустарникового (Salicornia fruticosa) и солероса европейского с участием лебеды прибрежной (Atriplex litoralis) и лебеды красив о плод ной (А. саlotheca), а в Австралии соответственно артрок-немума сарсазановидного, артрокнемума деревцевидного, солероса южного (Salicornia austraiis) и солероса деревцевидного (S. arbuscula), с участием лебеды болотной (Atriplex paludosa) и лебеды ножкоплодной (A. stipitata).
Многим, наверное, знаком вид мелких озер в степной и пустынной зоне нашей страны, окольцованных в начале лета изумрудно-зеленой полосой, а в конце лета — ярко-багряной или бордовой. Эта полоса образована однолетними суккулентными сведами, в массе развивающимися на сильно просоленной отлогой закраине озер, где из-за интенсивного испарения воды соль концентрируется в поверхностном слое почвы.
Произрастая в условиях постоянной почвенной и атмосферной сухости пустынь и степей или в условиях физиологической сухости засоленных почв и солончаков озерных и морских побережий, маревые обладают целым рядом специфических приспособлений. Это прежде всего очень высокое осмотическое давление в клетках их тканей. Клеточный сок у большинства маревых содержит такое большое количество растворенных солей, что зола многих из них издавна служила источником соды и поташа для местного населения и даже для промышленного использования. Многие маревые при этом являются и ярко выраженными суккулентами, как поташники, солеросы, сведы, галогетон (Halogeton), некоторые солянки, ежовники и др. У ряда маревых листья полностью редуцированы и ассимиляционную функцию выполняют зеленые ветви, как это особенно ярко выражено у саксаула. Этим достигается сильное сокращение испаряющей поверхности.
Сокращению испарения и защите от солнечного перегрева служит и хорошо развитый покров из различной формы волосков на листьях и молодых веточках, о чем уже говорилось выше. Например, у кокпека (Atriplex сапа), слагающего широко распространенные в пустынях Казахстана формации кокпечников, листья и молодые побеги серебристые от плотно покрывающих их чешуйчатых волосков, а у бассии пушистолистной (Bassia dasyphyllа), обычного в Центральной Азии рудераль-ного однолетника, все растение бело-пушистое от густых и длинных оттопыренных волосков.
Несомненно, как защита против иссушения выработалась и плотноподушковидная форма роста у ряда пустынных маревых, обитающих в крайне аридных условиях. Даже анатомическое строение осевых органов маревых, исключая трибу хруплявниковых, отличается большим своеобразием: сосудисто-волокнистые пучки у них расположены чередующимися кольцами, по типу однодольных древесных. Благодаря этому происходит быстрое одревеснение и образование перидермы даже у однолетников, и, например, крупный разросшийся экземпляр обыкновенного солероса легко принять за многолетнее растение. Эта особенность маревых также способствует их приспособлению к жизни в аридных условиях.
Поскольку маревые, за редким исключением, — растения открытых пространств, естественно, у них развито ветроопыление. Обеспечивается оно обычными способами — массой мелких открытых цветков, собранных в крупные и густые, чаще специализированные соцветия из тычиночных или пестичных цветков, или сильно выставляющимися тычинками и столбиками в случае обоеполых цветков, как у большинства солянок. По-видимому, как специфическое приспособление для ветроопы-ления надо рассматривать крупные пузыревидные придатки пыльников у некоторых солянок с закрытыми цветками, вибрация которых на ветру обеспечивает вытряхивание пыльцы из пыльников. Вместе с тем у ряда маревых (марь, свекла, ежовник) в цветках развит нектароносный подпестичный диск и отмечено насекомоопыление. Насекомые посещают цветки маревых ради обильной пыльцы, а также ради мясистых придатков пыльников.
Гораздо разнообразнее у маревых приспособления для рассеивания семян. Здесь прежде всего нужно упомянуть так называемое перекати-поле, когда высохшие кусты крупных однолетников с загнутыми внутрь ветвями приобретают шаровидную форму, отламываются у самой корневой шейки и свободно перекатываются ветром на большие расстояния, по пути рассеивая зрелые плоды. Чаще всего такие перекати-поле в степной полосе нашей страны принадлежат кураю (Salsola australis), самой распространенной однолетней солянке. Перекати-поле образуют также катун, или солянка холмовая (S. collina), кумарчик колючий (Agriophyllum pimgens, рис. 206), рогач, лебеда розовая (Atriplex rosea), верблюдка повислая (Corispermum declinatum), маръ остистая (Chenopodium aristatum) и многие другие.
Для распространения ветром предназначены все легко опадающие плоды с различной формы крыловидными придатками у большинства видов трибы солянковых (солянка, саксаульчик, саксаул, ежовник,галогетон и др.), а также у кохии и родственных родов. У терескена, пандерии (Panderia), лондезии, кириловии парусность плодов обеспечивается пучками и клубками длинных волосков, которыми снабжены окружающие плод прицветнички или листочки околоцветника. У многих видов лебеды этой же цели служат дискообразные или вееровидные легкие прицветнички.
У многих маревых плоды приспособлены для распространения их животными. Это осуществляется двумя путями: или они съедобные, или они цепляющиеся, либо накалывающиеся.
У съедобных плодов околоцветник или околоплодник становится сочным, мясистым и часто ярко окрашенным, а семена имеют плотную кожуру, не перевариваются в кишечном тракте животного и рассеиваются вместе с экскрементами. Чаще всего такие плоды распространяются птицами. Примером их могут служить ярко-красные соплодия жминды. У цепляющихся плодов маревых на твердеющем околоцветнике развиваются тонкие крючковидные выросты, легко цепляющиеся за шерсть животных, как у бассии иссополистной (Bassia hyssopifolia, рис. 205). Накалывающиеся плоды имеют острые твердые шипы, вонзающиеся в кожный покров или копыта животных. В качестве примера можно назвать колючие соплодия дикого шпината туркестанского (Spinacia turkesta-nica), плоды рогача (рис. 205), у которых шипы развиваются на сомкнутых прицветнич-ках, а также колючие плоды бассии пушисто-листной, околоцветник которых снабжен тонкими шипами.
В жизни человека маревые имеют большое значение. Важнейшим растением из них является свекла обыкновенная (Beta vulgaris), прежде всего ее разновидность — сахарная свекла. В странах умеренного пояса она является основным источником получения сахара. Получены и введены в культуру высокоурожайные новые гибридные, три- и тетраплоидные сорта с очень большим содержанием сахара в корнях — до 25%. Существенную роль в нашем повседневном пищевом рационе играет как овощ и столовая свекла с множеством ее сортов. В больших масштабах культивируется также кормовая свекла, корни и ботва которой — прекрасный корм для скота. В странах Западной и Южной Европы, в США, Японии, Индии широко распространена как овощ еще листовая (шпинатная) свекла, или мангольд, очень богатая витаминами А, В и С и минеральными солями.
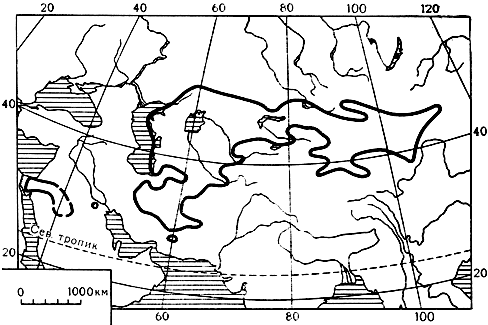
Карта 17. Ареал рода саксаул
История происхождения культурной свеклы довольно сложна и запутана, но несомненно, что родоначальницей культурной свеклы была дикая прибрежно-морская свекла (свекла морская — В. vulgaris subsp. maritima) и поныне произрастающая на побережьях Южной и Западной Европы (на севере до Западной Швеции). Первоначально в культуре появилась листовая (шпинатная) свекла, или мангольд. Она была известна как культурное растение уже в Вавилонии и впервые упоминается в письменных источниках в VIII в. до н. э. Позднее путем гибридизации ее с другими дикорастущими видами свеклы и селекции в культуре возникла столовая корнеплодная свекла. Кормовая свекла была выведена только в XVI в. в Германии и лишь в конце XVIII в. там же была открыта сахарная свекла. Однако способ получения сахара из нее и начало развития сахарной промышленности в Европе относится к 1800 г.
Всем известен как витаминоносный овощ шпинат огородный (Spinacia oleracea) — культурное растение, пришедшее к нам из Передней Азии в раннеисторическое время. Шпинат содержит значительное количество витаминов А, В и С, железа и фосфора; он очень богат белком (34% протеина от сухой массы; уступает лишь мясу) и поэтому является не только важным пищевым растением и диэтическим продуктом, но и укрепляющим средством при малокровии и истощении организма. Существует около двух десятков сортов культурного шпината, различающихся главным образом по форме (гладколистные и курчавые) и сочности листьев, а также по характеру олиственности стебля, времени цветения и т. д. В качестве шпината местное население в разных странах употребляет молодые листья многих дикорастущих маревых — мари белой, мари зеленой (Cheno-podium viride), мари городской (С. urbicum), мари прутьевидной (С. virgatum), лебеды садовой, лебеды копьевидной (Atriplex hastata), лебеды настоящей, галогетона, видов сведы, солянки, солероса.
В Перу, Чили, Боливии и Колумбии туземное население с первобытных времен возделывает в качестве хлебного растения киноа (Chenopodium quinoa) — культурный вид местного происхождения. Это чрезвычайно высокоурожайная неприхотливая культура, которую можно возделывать в горах до 4000 м высоты над уровнем моря, где уже никакие другие пищевые растения вызревать не могут. Зерно (семена) киноа исключительно питательно: оно содержит около 19% белка (протеина), свыше 47% крахмала, 4,8% жира и витамин В. Правда, семена киноа, как почти всех маревых, содержат сапонин, горькие и перед использованием в пищу требуют основательного вымачивания. Из-за горечи киноа как хлебное растение не привилось в европейских странах, но теперь как кормовое растение для скота его возделывают в США и Южной Африке, в Шри-Ланке. Аналогично в Гималаях туземное население в качестве хлебного растения возделывает марь белую и марь стенную (С. murale), а также использует их дикие заросли. Вообще в Европе и Азии в голодные годы население неизменно прибегало к мари и лебеде, как суррогату крупы и хлеба. Недаром ходила пословица: «Не то беда, что не хлеб, а лебеда — нет хуже беды, когда ни хлеба, ни лебеды». Калмыки и монголы употребляли с той же целью семена кумарчика.
Многие маревые являются лекарственными растениями, применяют их и в народной медицине и в официальной. Из них прежде всего следует назвать южноамериканские ароматичные мари — марь амброзиевидную (Chenopodium ambrosioides) и жарь противоглистную (С. anthelminticum), из семян которых получают эфирное масло, аскаридол, используемое как сильное глистогонное средство, а также в парфюмерии. Эти мари поэтому широко культивируются по всему свету, во многих странах одичали и стали почти космополитными сорными растениями. Сапонинсодержащие маревые имеют лекарственное действие как мочегонные, отхаркивающие, усиливающие сердечную деятельность и перистальтику кишечника, а также применяются против кожных заболеваний. Так, трава той же мари амброзиевидной известна как «мексиканский чай», используемый при простудных и желудочных заболеваниях. Подобным образом используют в разных странах и другие маревые. Как лекарственное растение, содержащее камфороподобное эфирное масло, применяют и камфоросму монпельенскую.
Распространенный в пустынях Северной Африки и у нас в Казахстане итсегек служит источником ценного препарата — инсектицида анабазин-сульфата, применяемого против сельскохозяйственных вредителей. Его производство налажено на базе естественных зарослей и плантаций итсегека в Южном Казахстане на специальном заводе в Чимкенте. Ряд маревых используют в народных промыслах в качестве превосходных красителей для тканей и кож (марь белая — красная краска для кож, марь зловонная — Chenopodium vulvaria — стойкая желтая краска, лебеда садовая — синяя, заменитель индиго), пищевых красителей (для вин, масла и т. п.), дубителей (бассия шерстистая — Bassia eriophora — и некоторые солянки), в качестве источников получения соды и поташа.
Велика роль маревых как кормовых пастбищных растений в аридных областях многих государств с развитым отгонным животноводством — в Китае и Монголии, Австралии и США, Алжире, Египте, Тунисе, в странах Ближнего Востока и Передней Азии, у нас в Средней Азии и Казахстане. Корну лака одно-колючковая (Cornulaca monacantha), солянка вонючая (Salsola baryosma), ежовник членистый, траганум (Traganum nudatum), бассия колючковая в Северной Африке и на Ближнем Востоке, биюргун, ежовник коротколистный (Anabasis brevifolia), изенъ (Kochia prostrata), терескен, кокпек, солянка воробьиная (Salsola passerina) и другие виды солянок в Казахстане, Средней и Центральной Азии — излюбленный корм для верблюдов во все времена года, а для овец, коз и крупного рогатого скота в осенне-зимнее время (молодые растения для них ядовиты). В пустынных районах Австралии кустарничковые виды лебеды и рагодии — единственный корм для овец.
В последнее время ряд однолетних и многолетних маревых все шире вводится в культуру для создания искусственных и улучшения естественных пастбищ в засушливых районах ряда стран. Всем известный саксаул — легендарное дерево песчаных пустынь — не столь значим как топливо, сколько как закрепитель песков и основа долговременных пастбищ в песчаных пустынях Средней Азии. С этой целью его ежегодно высевают с самолетов на тысячах гектаров сыпучих песков, так же как и крупнокустарниковые солянки — солянку Рихтера (Salsola richteri, табл. 59) и солянку Палецкого (S. paletzkiana).
Все интенсивнее вводятся в культуру кустарниковые и полукустарниковые маревые как засухо- и солеустойчивые кормовые растения для пустынных пастбищ. В Средней Азии в больших масштабах культивируется изень, уже получившая широкое признание как пастбищное кормовое растение и интродуцированная в Аргентину, США, Египет. Австралийские виды лебеды — лебеда монетная (Atriplex nummularia), лебеда карликовая (A. pnmilio), лебеда полуприцветниковая (A. semibracteata) и средиземноморская лебеда настоящая (A. halimus) — культивируются в Австралии, Израиле, Эфиопии, Южной Африке, США. С этой же целью в США испытываются и вводятся в культуру местные виды лебеды (Atriplex canescens, A. halimoides, A. leptocarpa, A. vesicaria). Особенно высокие кормовые достоинства обнаружены у австралийской лебеды монетной, и она интродуцирована теперь в Северную и Южную Африку, Южную Америку, Израиль. В Тунисе местная лебеда настоящая выращивается как кормовое сенокосное растение на тысячах гектаров и ее посевы расширяются с каждым годом.
Среди маревых мало растений, используемых в качестве декоративных. Пожалуй, наибольшее распространение получила разновидность обычной кохии веничной (Kochia scoparia var. trichophylla) с плотным кустом и яркой зеленью для клумб и как горшечное комнатное растение. В качестве ампельного настенного декоративного растения используется травянистая лиана габлиция тамусовидная. Далее можно упомянуть культивируемые садовые пестролистные формы мари багрянистой (Chenopodmm purpurascens) и лебеды садовой, а также киноа, имеющую крупные малиново-красные соцветия, и жминду с ярко-красными соплодиями.
Вначале уже упоминалось, что ряд маревых, особенно марь и лебеда, входят в число самых распространенных и зловредных сорняков полей, садов и огородов — сорняков, трудно искоренимых. Этому способствуют две биологические особенности мари и лебеды: колоссальное количество производимых одним растением семян и их разнокачественность. Например, на плодородной почве обыкновенная марь белая развивает куст высотой до 2 м и в обхвате до 4 м и приносит более 100 000 семян. При этом на одном и том же растении образуются семена трех видов: 1) крупные, плоские, коричневые, прорастающие при высеве через 3 дня; 2) более мелкие, с толстой оболочкой, линзовидные черные, прорастающие на второй год, и 3) очень мелкие, почти круглые, черные, прорастающие при благоприятных условиях лишь на третий год, но сохраняющие всхожесть много лет. К тому же марь — растение необыкновенно выносливое и распространено по всему свету от полярного круга до тропиков; проростки ее легко выдерживают весенние заморозки до -6 °С, быстро растут. Подобными качествами обладают и многие другие сорные маревые, поэтому бороться с ними очень трудно.
Источник статьи: http://plantlife.ru/books/item/f00/s00/z0000028/st136.shtml
