Экология СПРАВОЧНИК
Различные типы леса представлены то хвойными, не сбрасывающими на зиму хвою, то лиственными, сбрасывающими на зиму листву. В одних типах леса — породы, имеющие широкую густую крону, которая является более надежным от перегрева экраном (липа). В других породы имеют узкую крону (ель, пихта, кипарисы, тополь пирамидальный). В различных типах леса выделяются разные фитонциды, притом в неодинаковых количествах (сосна в чистом сосновом бору на песках выделяет больше терпенов, в сложном бору, со вторым ярусом из ели меньше терпенов): липа выделяется по сравнению, например, с осиной ароматом своих цветов, черемуха — запахом своих цветов и специфическим запахом листьев и ветвей при растирании первых и надламывании вторых. Распускание весной хвои лиственницы отличает лесные сообщества с этой породой особым нежным запахом, которого нет на участках с господством ели.[ . ]
Широколиственные сообщества представлены дубом, липой, ильмом, осиной. В травяном покрове основным растением является сныть, к которой в более северных районах могут примешиваться чина, ясменник, копытень, звездчатка, а в более южных—вейник, миндаль-бобовник и др. Для подлеска широколиственных лесов характерны орешник, бересклет, рябина. Большинство современных лиственных лесов являются вторичными (березовые, осиновые, липовые), возникшими на месте вырубок коренных хвойных и части широколиственных лесов.[ . ]
В пределах профиля нами отмечено 22 растительных сообщества (растительность скальных выходов, каменных россыпей и пойм, подгольцо-вые редколесья, лиственные и темнохвойные леса, высокотравья, растительность тропы и туристской стоянки «Белый ключ»).[ . ]
Зольность опада (с учетом надземной части и корней) в лугово-степных сообществах составляет 7—8 %, в хвойных лесах — 0,7—1,7 % и в лиственных — 1,6—7,5 % (Базилевич, 1962). Содержание азота также самое высокое в опа-де лугово-степных сообществ (1—1,4 %).[ . ]
Рассмотрим каждый из уровней. Процент площадей, способных поддерживать сообщества в состоянии климатического климакса, различен для разных областей. Но поскольку стратегия развития любой экосистемы состоит в достижении климатического климакса, то главными наземными экосистемами можно считать биомы. Они легко выделяются, в частности, по климатической кпимаксной растительности (рис. 13.2). По этому показателю можно выделить тундру, северный хвойный лес (тайгу), лиственный лес умеренной зоны, степь, пустыню, тропический дождевой лес и т. д.[ . ]
Давно замечено, что восстановление растительности на ете вырубленного елового леса происходит не сразу, а через ряд стадий каждая из которых характеризуется определенным типом сообщества, впоследствии сменяемого другим. Со временем он» догоняют в росте лиственные породы и возникает та-им образом смешанный лес.[ . ]
Необходимо отметить, что, несмотря на сильные изменения состава и структуры лесных сообществ в разных стадиях возрастного развития, диагнозирование их принадлежности к определенному типу леса, отличающемуся своеобразным лесообразовательным процессом, вполне возможно, но для этого необходимы специальные исследования не только фитоценотического состава, но и других компонентов биогеоценоза, охватывающие все стадии развития насаждений. В условиях Литвы такие исследования проведены в елово-лиственных лесах, но пока еще недостаточно изучены сосновые леса.[ . ]
| Можно обнаружить иерархию местообитаний, как бы вставленных друг в друга | 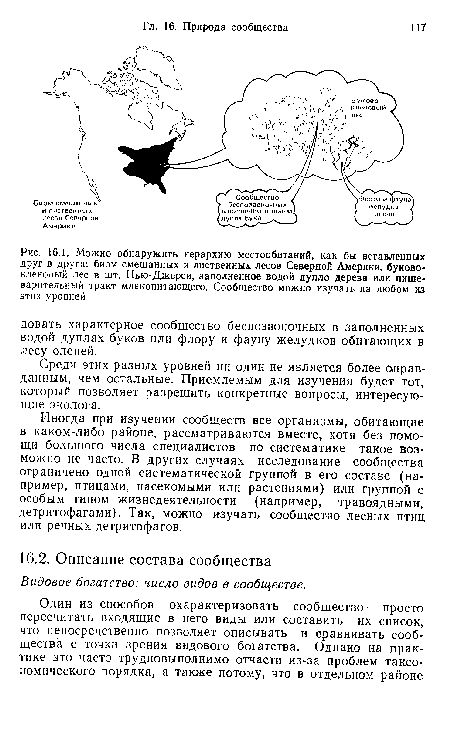 |
С формированием молодняка начинается сукцессионный процесс, в результате которого растительность вырубок сменяется живым напочвенным покровом лесного сообщества. Изучение этого процесса показало, что от вырубки до спелого леса можно выделить 5 стадий, различающихся по густоте и составу растительного покрова, а также и по другим компонентам биогеоценоза [4, 3,2]. Изменение проективного покрытия и состава живого напочвенного покрова в процессе возрастного развития елово-лиственных древостоев кислично-широко-травного типа леса показано на рис. 2.[ . ]
Деятельность редуцентов позволяет всему годичному опаду (опадающих листьев, цветов, плодов, трав и т. п.) органических веществ полностью разложиться в тропиках во влажных дождевых лесах за 1—2 года, в умеренной зоне — в лиственных лесах за 2—4 года, в хвойных лесах за 4—5 лет. В тундре процесс разложения продолжается десятками лет. Во влажных условиях возрастает способность грибов и некоторых бактерий образовывать громадные количества спор (респро-дуктивных клеток). Эти микроскопические частицы переносятся воздушными потоками в атмосфере на весьма значительные расстояния, что позволяет им распространяться повсеместно и давать жизнеспособное потомство на любом пространстве при наличии оптимальных условий жизнедеятельности. Активность деятельности редуцентов серьезным образом зависит от факторов среды: температуры, влажности, инсоляции и др. Интенсивность процессов редуцирования определяется также и биологическими процессами, в частности продуктивностью растительных сообществ, активностью консументов, а на данном этапе и антропогенными факторами.[ . ]
Величина p-разнообразия, определенная с помощью коэффициента Жаккара (К), зависит в данном случае от изменения высоты в значительно меньшей степени, чем от антропогенной нагрузки и условий увлажнения. Из числа естественных сообществ, высокое своеобразие отличает только пойменную растительность (ручья — К = 0.0-0.22, и р. Б. Тесьмы — К = 0.0-0.23), тогда как для растительности всех описанных антропогенных местообитаний характерно высокое своеобразие (растительность тропы — К = 0.10-0.32, туристской стоянки — К = 0.0-0.20, опушки леса около стоянки — К = 0.0-0.18). Наибольшим ценотическим сходством отличаются описанные участки лиственных лесов.[ . ]
Примером вторичных демутационных сукцессий может служить восстановление климаксового лесного биоценоза после пожаров (а в наше время и вырубок). В таежной зоне Евразии при появлении открытого пространства на месте еловых лесов в результате пожара или сплошной рубки коренным образом меняется режим освещения, температуры, влажности и других факторов. Эти изменения неблагоприятны для таежных видов растений и животных, развитие которых угнетается. Зато на освещенных, относительно сухих и хорошо прогреваемых местах формируется временное одноярусное сообщество из светолюбивых трав. Первыми на стадии открытой вырубки развиваются виды с легко распространяемыми семенами: вейник, Иван-чай и др. Позднее начинается лесовозобновление: прорастают светолюбивые лиственные породы (осины, березы, ивы и др.) и кустарники. Одновременно формируется связанный с лугово-кустарниковой растительностью комплекс животного населения: многочисленные и разнообразные насекомые и другие беспозвоночные, грызуны, исходно связанные с лугами и лесными полянами; развитие ягодных кустарников привлекает большое число видов птиц, а наличие хорошо прогреваемых мест — рептилий и т. д. Формируется богатое и разнообразное сообщество зарастающей вырубки.[ . ]
Флора этой провинции, несмотря на ее размеры, содержит, вероятно, лишь немногим более 1500 видов. Но в ней имеются эндемичные роды (злак Тшюипа, бурачниковое апэсЬа-ше11а и др.) и много эндемичных видов. В растительном покрове доминируют степные сообщества, кобрезиевые «ковры» и осоковые болота. Некоторые авторы называют Центральный Тянь-Шань страной злаков и осок. Что же касается древесно-кустарниковой растительности, то она представлена здесь фрагментарно и занимает небольшие площади. В глубине Центрального Тянь-Шаня, особенно на Терскей-Алатау и Кунгей-Алатау, имеются отдельные, обычно небольшие, островки еловых лесов. Встречаются также лиственные насаждения, состоящие из тополя, березы, видов жимолости, изредка дикого абрикоса и др.[ . ]
Процессы формирования растительности вырубок изучаются нами уже почти 20 лет, поэтому некоторые данные можно представить как в статике, так и в динамике. Следуя концепции типологии вырубок И. С. Мелехова, по данным рекогносцировочного исследования вырубок еловых и елово-лиственных лесов Литвы определено примерное распределение вырубок по доминантам растительного покрова. Эти данные (табл. 1) показывают, что большинство доминант сообществ растительности вырубок являются растениями широкого экологического диапазона и встречаются в местопроизрастаниях нескольких типов. Кроме того, в местах произрастания одного типа леса могут доминировать разные растения; их распространение зависит от причин, уже упомянутых выше, а также от биологических свойств самих растений. В основном от этих свойств зависит изменение состава растительности на вырубках в связи с возрастом последних. Иллюстрацией этих положений может быть динамика растительного покрова вырубок двух типов елово-лиственных лесов в западной части Литвы (Таурагский леспромхоз) , показанная на рис. 1.[ . ]
Термин «экосистема» введен в экологию английским ботаником А. Тенсли (1935). Понятие экосистемы не ограничивается какими-то признаками ранга, размера, сложности или происхождения. Поэтому оно приложимо как к относительно простым искусственным (аквариум, теплица, пшеничное поле, обитаемый космический корабль), так и к сложным естественным комплексам организмов и среды их обитания (озеро, лес, океан, экосфера). Различают водные и наземные экосистемы. Все они образуют на поверхности планеты густую пеструю мозаику. При этом в одной природной зоне встречается множество сходных экосистем — или слитых в однородные комплексы, или разделенных другими экосистемами. Например, участки лиственных лесов, перемежающиеся хвойными лесами, или болота среди лесов и т.п. В каждой локальной наземной экосистеме есть абиотический компонент — биотоп, или экотоп, — участок с одинаковыми ландшафтными, климатическими, почвенными условиями и биотический компонент — сообщество, или биоценоз, — совокупность всех живых организмов, населяющих данный биотоп. Биотоп является общим местообитанием для всех членов сообщества. Биоценозы состоят из представителей многих видов растений, животных и микроорганизмов. Практически каждый вид в биоценозе представлен многими особями разного пола и возраста. Они образуют популяцию (или часть популяции) данного вида в экосистеме.[ . ]
Второй пример иллюстрирует изменения в видовом составе и плотности гнездящихся птиц (табл. 39), параллельные изменениям доминирующих растений, во вторичной сукцессии, начавшейся на заброшенных пахотных землях в гористой местности на юго-востоке США (фиг. 124). Обратите внимание, что самые значительные изменения в популяциях птиц возникают при изменении жизненных форм доминирующих растений (травы, кустарники, сосна и лиственные породы). Ни один из видов растений или птиц не способен процветать на протяжении всей сукцессии, максимумы разных видов приходятся на разные точки временной шкалы. Хотя растения играют наиболее важную роль в качестве организмов, вызывающих изменения, это отнюдь не означает, что птицы играют в сообществе совершенно пассивную роль: распространение в новые области семян главных доминирующих растений из числа ку-стяоников и лиственных деревьев целиком зависит от птиц и других животных. Терминальная, или климаксная, стадия — лес из дуба и гикори, а не буково-кленовый лес, как в предыдущем примере, что связано с региональными различиями в климате. Б), ускоряя таким образом их смену астрами. Как уже говорилось в предыдущем разделе, возможно, что такие «химически индуцированные» сукцессии встречаются довольно часто. Подводя итоги, следует сказать, что, хотя климат и другие физические факторы контролируют состав сообществ и определяют климакс, важную роль в развитии сукцессии играют сами сообщества.[ . ]
Провинция характеризуется богатой и своеобразной флорой с очень высоким видовым эндемизмом. Имеются также эндемичные секции и подроды, более двух десятков эндемичных родов. Особенно много эндемичных таксонов в семействах сложноцветных, зонтичных, бурачниковых, крестоцветных, маревых, бобовых, губоцветных, лилейных, свинчатковых, мареновых и норичниковых. Характерны пустынные и полупустынные формации, горные степи, часто с участием колючих подушкообразных кустарничков (особенно колючий эспарцет Оп-obrychis cornuta и виды Acantholimon), сообщества ксерофильных и жестколистных кустарников и кустарничков и можжевеловые и лиственные редколесья. Во многих местах распространены ксерофильные дубовые леса.[ . ]
Источник