Регуляция глюконеогенеза.
Важным моментом в регуляции глюконеогенеза является реакция, катализируемая пируваткарбоксилазой. Роль положительного аллостерического модулятора этого фермента выполняет ацетил-КоА. В отсутствие ацетил-КоА фермент почти полностью лишен активности. Когда же в клетке накапливается митохондриальный ацетил-КоА, биосинтез глюкозы из пирувата усиливается. Известно, что ацетил-КоА одновременно является отрицательным модулятором пируватдегидрогеназного комплекса. Следовательно, накопление ацетил-КоА замедляет окислительное декарбоксилирование пирувата, что также способствует превращению в глюкозу.
Вторым важным моментом в регуляции глюконеогенеза является реакция, катализируемая фруктозо-1,6-бифосфатазой — ферментом, который ингибируется АМФ. Противоположное действие АМФ оказывает на фосфофруктокиназу, т.е. для этого фермента он является аллостерическим активатором. При низкой концентрации АМФ и высоком уровне АТФ происходит стимуляция глюконеогенеза. Напротив, когда величина отношения АТФ/АМФ низка, в клетке наблюдается расщепление глюкозы.
В 1980 г. группой бельгийских исследователей (Г.Херс и соавт.) в ткани печени был открыт фруктозо-2,6-бисфосфат, который является мощным регулятором активности двух вышеперечисленных ферментов:
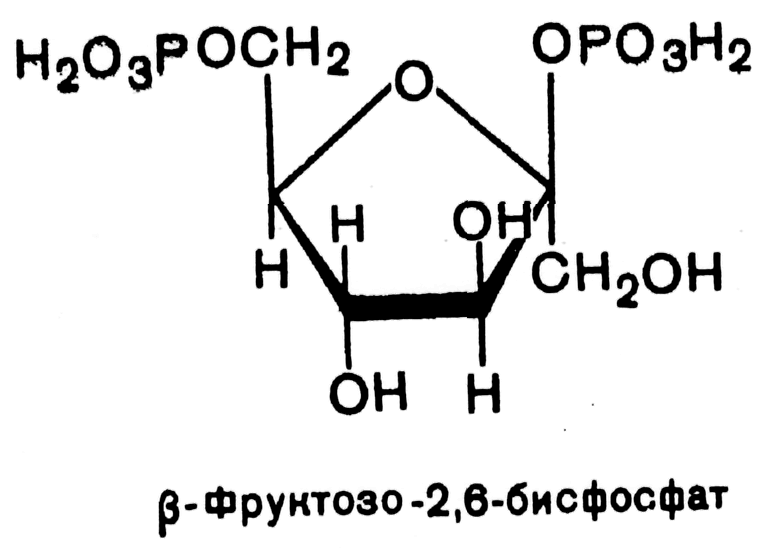
Фруктозо-2,6-бисфосфат активирует фосфофруктокиназу и ингибирует фруктозо-1,6-бифосфатазу. Повышение в клетке уровня фруктозо-2,6-бисфосфата способствует усилению гликолиза и уменьшению скорости глюконеогенеза.
При снижении концентрации фруктозо-2,6-бисфосфата отмечается обратная картина.
Установлено, что биосинтез фруктозо-2,6-бисфосфата происходит из фруктозо-6-фосфата при участии АТФ, а распадается он на фруктозо-6-фосфат и неорганический фосфат. Биосинтез и распад фруктозо-2,6- бисфосфата катализируется одним и тем же ферментом, т.е. данный фермент бифункционален, он обладает и фосфокиназной и фосфатазной активностями:

АТФ АДФ


Фруктозо-6-фосфат фосфокиназное действие Фруктозо-2,6-бисфосфат

Показано также, что бифункциональный фермент в свою очередь регулируется путем цАМФ-зависимого фосфорилирования. Фосфорилирование приводит к увеличению фосфатазной активности и снижению фосфокиназной активности бифункционального фермента. Этот механизм объясняет быстрое действие гормонов, в частности глюкагона, на уровень фруктозо-2,6-бисфосфата в клетке.
Активность бифункционального фермента регулируется также и некоторыми метаболитами, среди которых наибольшее значение имеет глицерол-3-фосфат. Действие глицерол-3-фосфата на фермент по своей направленности аналогично эффекту, который наблюдается при его фосфорилировании с помощью цАМФ-зависимых протеинкиназ.
В настоящее время фруктозо-2,6-бисфосфат, помимо печени, обнаружен и в других органах и тканях животных, а также у растений и микроорганизмов.
Показано, что глюконеогенез может регулироваться и непрямым путем, т.е. через изменение активности фермента, непосредственно участвующего в синтезе глюкозы. Так, установлено, что фермент гликолиза пируваткиназа существует в двух формах: L и М. Форма L (от англ. liver — печень) преобладает в тканях, способных к глюконеогенезу. Эта форма ингибируется избытком АТФ и некоторыми аминокислотами, в частности анилином. М-форма (от англ. Muscle — мышца) такой регуляции не подвержена. В условиях достаточного обеспечения клетки энергией имеет место ингибирование L-формы пируваткиназы, и как следствие этого, гликолиз замедляется и создаются условия, благоприятствующие глюконеогенезу.
Источник
Взаимопревращения моносахаридов
Конечный продукт цикла Кальвина фруктозо-6-фосфат путём изомеризации очень легко превращается в глюкозо-6-фосфат под действием фермента глюкозофосфатизомеразы:
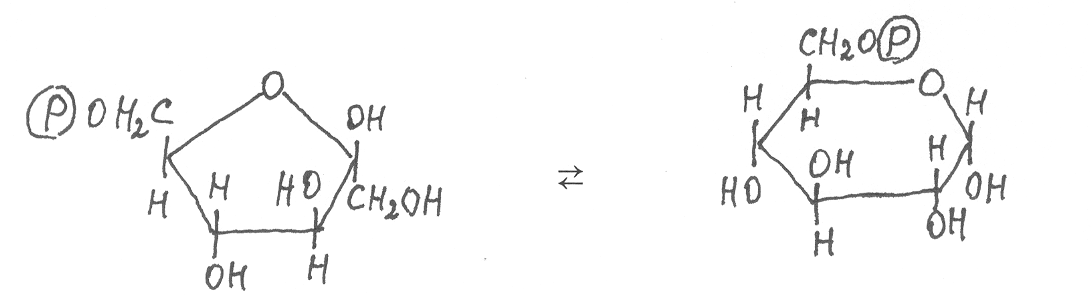
фруктозо-6-фосфат глюкозо-6-фосфат
Вследствие того, что реакции изомеризации сопровождаются небольшим изменением свободной энергии, они в большинстве своём обратимы.
Под действием фермента маннозофосфатизомеразы фруктозо-6-фосфат изомеризуется в маннозо-6-фосфат:

фрукто-6-фосфат маннозо-6-фосфат

Врезультате действия соответствующих мутаз, осуществляющих внутримолекулярный перенос фосфатной группы, из глюкозо-6-фосфата образуется глюкозо-1-фосфат, а из маннозо-6-фосфата – маннозо-1-фосфат:

маннозо-6-фосфат маннозо-1-фосфат
В нефотосинтезирующих клетках растений и у гетеротрофных организмов фруктозо-6-фосфат и глюкозо-6-фосфат включаются в реакции гликолиза или пентозофосфатного цикла. В ходе реакций гликолиза фосфорилированная гексоза расщепляется на две триозы – 3-фосфогли-цериновый альдегид и фосфодиоксиацетон, из которых при обращении реакций гликолиза может снова синтезироваться гексоза. В пентозофосфатном цикле образуется примерно такой же набор фосфорнокислых производных моносахаридов, как и в цикле Кальвина. Таким образом, мы видим, что в гликолитических реакциях осуществляются взаимные превращения гексоз и триоз на уровне их фосфорнокислых эфиров, а в пентозофосфатном цикле и цикле Кальвина происходят взаимные превращения фосфорилированных форм триоз, пентоз, эритрозы, глюкозы и фруктозы.
Включение свободных моносахаридов в различные реакции их превращений осуществляется путем фосфорилирования. В качестве донора фосфатных групп для реакций фосфорилирования чаще всего используется АТФ. Ферменты, катализирующие перенос фосфатных групп от АТФ на моносахариды, как мы уже знаем, называют киназами.
Фосфорилирование глюкозы и маннозы катализирует фермент гексокиназа с образованием глюкозо-6-фосфата и маннозо-6-фосфата.
По аналогичному механизму фруктокиназа катализирует образование фруктозо-1-фосфата из фруктозы, а фермент галактокиназа синтезирует галактозо-1-фосфат из галактозы.
Образование свободных моносахаридов из их фосфорилированных форм катализируют ферменты гидролитического типа – фосфатазы, которые отщепляют от фосфатов моносахаридов остатки фосфорной кислоты. Гидролиз глюкозо-6-фосфата происходит с участием глюкозо-6-фосфатазы, галактозо-1-фосфата – под действием галактозо-1-фосфатазы, фруктозо-6-фосфата – фруктозо-6-фосфатазы и т.д.

глюкозо-6-фосфат глюкоза
Значительно сложнее происходят взаимопревращения галактозы и других моносахаридов. В этих реакциях в качестве промежуточных продуктов образуются нуклеозиддифосфатпроизводные соответсвующих моносаха-ридов. На первом этапе таких превращений галактоза фосфорилируется с участием фермента галактокиназы, в результате образуется галактозо-1-фосфат:
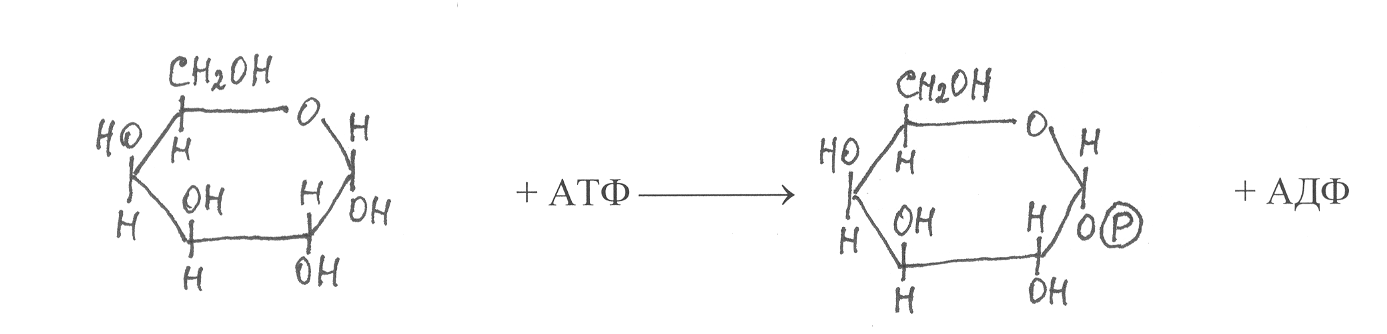
галактоза галактозо-1-фосфат

На следующей стадии галактозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ). Эту реакцию катализирует ферментгалактозо-1-фосфатуридилилтрансфераза, под действием которого обра-
зуется нуклеотидное производное галактозы – уридиндифосфат-галактоза (УДФ-галактоза) и пирофосфат.
В дальнейшем УДФ-галактоза изомеризуется в УДФ-глюкозу под действием специфической НАД-зависимой 4-эпимеразы:
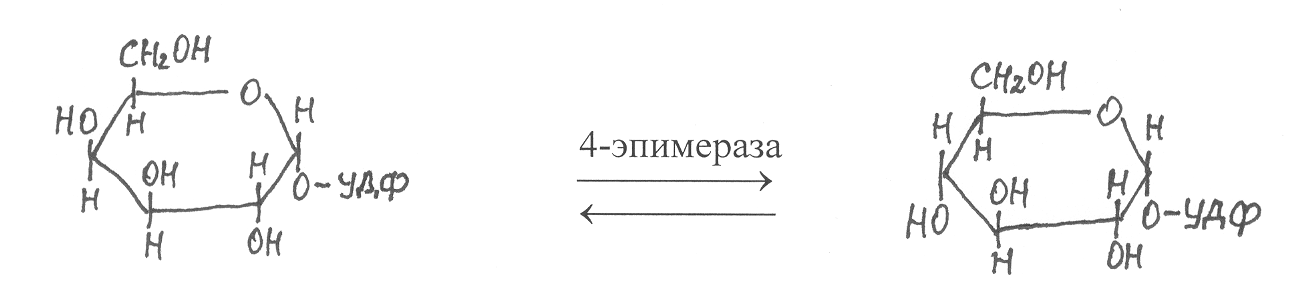
УДФ-галактоза УДФ-глюкоза
После гидролитического расщепления УДФ-глюкоза распадается на два продукта – глюкозо-1-фосфат и уридинмонофосфат (УМФ):

УДФ-глюкоза глюкозо-1-фосфат
Глюкозо-1-фосфат может далее изомеризоваться в глюкозо-6-фосфат, а глюкозо-6-фосфат – во фруктозо-6-фосфат. Таким образом, посредством указанных реакций галактоза может превращаться во фруктозо-6-фосфат, который включается в реакции дыхания, или в глюкозо-6-фосфат, способный превращаться в продукты пентозофосфатного цикла.
Возможен также синтез галактозо-1-фосфата из глюкозо-1-фосфата, так как в клетках организмов содержится фермент глюкозо-1-фосфатури-дилилтрансфераза, катализирующий образование УДФ-глюкозы из глюкозо-1-фосфата и УТФ:
Затем УДФ-глюкоза под действием 4-эпимеразы изомеризуется в УДФ-галактозу, при гидролизе которой образуется галактозо-1-фосфат:
УДФ-галактоза + Н2О¾¾®галактозо-1-фосфат + УМФ
Взаимопревращения гексоз и пентоз осуществляются в пентозофосфатном цикле и цикле Кальвина. Важное значение для этих реакций имеют ферменты транскетолаза и трансальдолаза, а в пентозо-фосфатном цикле – ещё и фермент фосфоглюконатдегидрогеназа, ката-лизирующий окислительное декарбоксилирование 6-фосфоглюконовой кислоты с образованием рибулозо-5-фосфата. Этот фермент фактически осуществляет превращение гексозы в пентозу. Во взаимных превращениях пентоз также участвуют ферменты рибулозо-фосфатэпимераза и рибозофосфатизомераза, поддерживающие динамическое равновесие между рибулозо-5-фосфатом, с одной стороны, и ксилулозо-5-фосфатом и рибозо-5-фосфатом, с другой стороны.
Ксилоза и арабиноза синтезируются также из гексоз, но другим путём. При этом в качестве промежуточных продуктов образуются нуклеотидные производные глюкуроновой и галактуроновой кислот. На первом этапе осуществляется ситез УДФ-глюкозы из глюкозо-1-фосфата и
УТФ, а затем под действием фермента УДФ-глюкозодегидрогеназы(1.1.1.22) УДФ-глюкоза окисляется в УДФ-глюкуроновую кислоту:
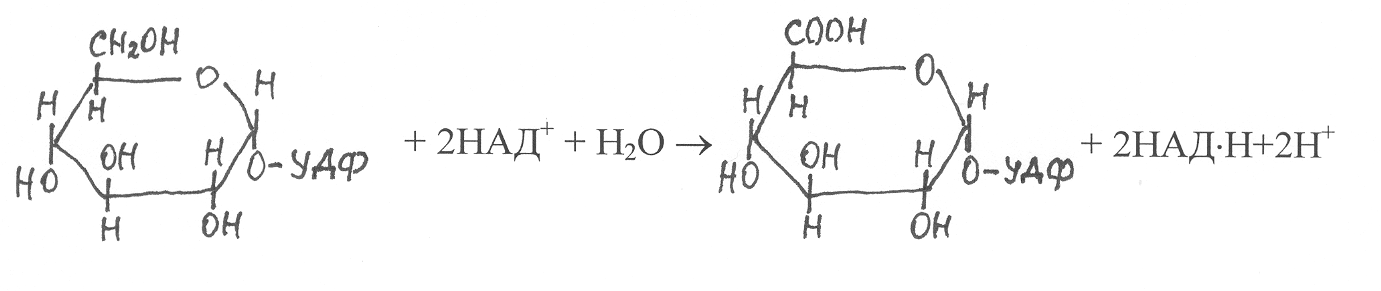
УДФ-глюкоза УДФ-глюкуроновая

Затем УДФ-глюкуроновая кислота подвергается декарбоксили-рованию и превращению в пиранозную форму УДФ-ксилозы:
Полученная таким путём УДФ-ксилоза используется в процессе синтеза ксиланов.

По аналогичному механизму осуществляется синтез УДФ-арабинозы из УДФ-галактозы, при этом в качестве промежуточного продукта образуется УДФ-галактуроновая кислота. УДФ-арабиноза так же, как и УДФ-ксилоза, не накапливается в растительных тканях, а используется для синтеза арабанов. Кроме того, возможны взаимные превращения УДФ-глюкуроновой и УДФ-галактуроновой кислот, а также пираназных форм УДФ-ксилозы и УДФ-арабинозы под действием соответствующих 4-эпимераз.
УДФ-глюкуроновая кислота УДФ-галактуроновая кислота
УДФ-галактуроновая кислота является основным источником галактуроновой кислоты для синтеза пектиновых веществ, а УДФ-глюкуроновая кислота участвует в синтезе ксиланов (в качестве ответвлений), полиуренидов, аскорбиновой кислоты.
Превращение моносахаридов в спирты. У растений, грибов и водорослей важную роль в углеводном обмене играют спирты – сорбит, маннит, дульцит.
Дульцита много содержится в листьях ряда растений, особенно в листьях бересклета. Сорбита много накапливается в плодах и ягодах. Очень много маннита в грибах и водорослях, а также в некоторых растительных продуктах. Синтез этих спиртов осуществляется в результате восстановления соответствующих моносахаридов. Наиболее хорошо изучена реакция образования маннита из фруктозы. Эту реакцию катализирует фермент маннитолдегидрогеназа (1.1.1.138):
Источник