- Бескислородное окисление глюкозы включает два этапа
- Гликолиз
- Первый этап гликолиза
- Второй этап гликолиза
- БИОХИМИЯ — Л. Страйер — 1984
- ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
- ГЛАВА 19. ФОТОСИНТЕЗ
- 19.15. СO2 реагирует с рибулозобисфосфатом с образованием двух фосфоглицератов
- 19.16. Образование фруктозо-6-фосфата и регенерирование рибулозо-1,5-бисфосфата
- 19.17. Три АТР и два NADPH доводят СO2 до уровня гексозы
Бескислородное окисление глюкозы включает два этапа
В анаэробном процессе пировиноградная кислота восстанавливается до молочной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат далее ни во что не превращается, единственная возможность утилизовать лактат – это окислить его обратно в пируват.
Многие клетки организма способны к анаэробному окислению глюкозы. Для эритроцитов он является единственным источником энергии. Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсивную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спорта. Вне физических нагрузок бескислородное окисление глюкозы в клетках усиливается при гипоксии – при различного рода анемиях, при нарушении кровообращения в тканях независимо от причины.
Гликолиз
Анаэробное превращение глюкозы локализуется в цитозоле и включает два этапа из 11 ферментативных реакций.
Первый этап гликолиза
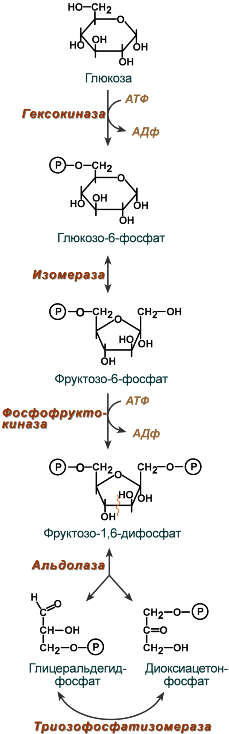
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой .
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент глюкозофосфат-изомераза ). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат- альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы . Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
- при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза,
- при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез глицеррол-3-фосфата для образования фосфатидной кислоты и далее тривцилглицеролов.
Второй этап гликолиза
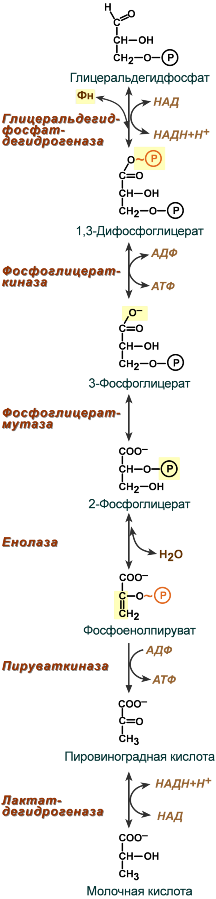
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа ) – окисление глицеральдегидфосфата до кислоты и присоединение к ней фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа ) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (использование энергии электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ с образованием пировиноградной кислоты и АТФ.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая – образование молочной кислоты из пирувата под действием лактатдегидрогеназы . Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях.
| У плода и детей первых месяцев жизни преобладает анаэробный распад глюкозы, в связи с чем концентрация молочной кислоты в крови у них выше чем у взрослых. |
| При наличии кислорода пировиноградная кислота переходит в митохондрию и превращается в ацетил-S-КоА. |
Источник
БИОХИМИЯ — Л. Страйер — 1984
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 19. ФОТОСИНТЕЗ
19.15. СO2 реагирует с рибулозобисфосфатом с образованием двух фосфоглицератов
Радиохроматограмма, получаемая после 60 с освещения, оказалась настолько сложной (рис, 19.19), что на ней невозможно было отличить самый ранний промежуточный продукт в процессе фиксации СО2. Однако после освещения в течение всего лишь 5 с картина была значительно проще: на хроматограмме присутствовало только одно явное радиоактивное пятно, которое оказалось 3-фосфоглицератом.
Рис. 19.18. Электронная микрофотография мембран тилакоидов шпината, полученная методом замораживания — скалывания. Видна правильная решетка трансмембранных частиц

Рис. 19.19. Радиохроматограммы освещенных суспензий водоросли через 5 и 60 с после введения в систему СO2

Образование 3-фосфоглицерата в качестве первого обнаруживаемого радиоактивного промежуточного продукта наводило на мысль, что акцептором СО2 служит двухуглеродное соединение. Оказалось, что это не так. Действительная последовательность реакций гораздо сложнее:

Молекула СО2 конденсируется с рибулозо-1,5-бисдифосфатом с образованием короткоживущего шестиуглеродного соединения, которое быстро гидролизуется до двух молекул 3-фосфоглицерата. Суммарная реакция носит высокоэкзергонический характер (∆G 0 = 12,4 ккал/моль).

Реакция катализируется рибулозо-1,5-бисфосфат—карбоксилазой, которая локализована на обращенной к строме поверхности мембран тилакоидов. Этим ферментом очень богаты хлоропласты; на его долю приходится более 16% общего содержания в них белка. Рибулозо-1,5-бисфосфат—карбоксилаза, вероятно, является белком, содержащимся в биосфере в наибольшем количестве. Он состоит из восьми больших (55 кДа) и восьми малых (15 кДа) субъединиц, расположенных в виде двух слоев (рис. 19.20). Большие субъединицы каталитически активны и в отсутствие малых субъединиц, которые выполняют регуляторную роль. Фермент является в одно и то же время и оксигеназой (разд. 19.20), и карбоксилазой.
Рис. 19.20. Схематическое изображение структуры рибулозо-1,5-бис- фосфат—карбоксилазы

19.16. Образование фруктозо-6-фосфата и регенерирование рибулозо-1,5-бисфосфата
Этапы превращения 3-фосфоглицерата во фруктозо-6-фосфат (рис. 19.21) подобны таковым в пути глюконеогенеза (разд. 15.13) с той лишь разницей, что глицеральдегид-3- фосфат—дегидрогеназа хлоропластов специфична в отношении NADPH, а не NАDН. В результате этих реакций СO2 переходит на уровень гексозы. Остается лишь регенерировать рибулозобисфосфат, акцептор СO2 на первой темновой стадии. Проблема сводится к образованию пятиуглеродного сахара из шестиуглеродного и трехуглеродного сахаров. Оно осуществляется в ходе реакций, катализируемых транскетолазой и альдолазой. Транскетолаза участвует также в превращениях по пентозофосфатному пути (разд. 15.4).

Напомним, что транскетолаза, фермент, использующий в качестве простетической группы тиаминпирофосфат, переносит двухуглеродный фрагмент (СН2ОН—СО—) с кетозы на альдозу. Аль- долаза осуществляет альдольную конденсацию между дигидроксиацетонфосфатом и альдегидом. Фермент этот высокоспецифичен для дигидроксиацетонфосфата, но реагирует с множеством альдегидов. К специфическим реакциям, катализируемым транскетолазой и альдолазой в цикле Кальвина, относятся следующие:

Рис. 19.21. Путь превращения 3-фосфо- глицерата во фруктозо-6-фосфат в хлоропластах

Для темновых реакций фотосинтеза требуются четыре дополнительных фермента. Один из них, фосфатаза, гидролизует седо- гептулозо-1,7-бисфосфат до седогептулозо-7-фосфата. Второй, фосфопентозо-эпиме- раза, превращает ксилулозо-5-фосфат в рибулозо-5-фосфат. Третий, фосфопентозо-изомераза, превращает рибозо-5-фосфат в рибулозо-5-фосфат. Напомним, что второй и третий ферменты участвуют также в превращениях по пентозофосфатному пути (разд. 15.4). Суммируя вышеуказанные реакции, получаем
Фруктозо-6-фосфат + 2-глицеральдегид-З- фосфат + Дигидроксиацетонфосфат → 3 Рибулозо-5-фосфат.
Наконец, четвертый фермент, фосфорибулозокиназа, катализирует фосфорилирование рибулозо-5-фосфата с регенерированием ри- булозо-1,5-бисфосфата, акцептора СO2.
Рибулозо-5-фосфат + АТР → Рибулозо-1,5-бисфосфат + ADP + Н + .
Эта группа реакций носит название цикла Кальвина (рис. 19.22).
Рис. 19.22. Цикл Кальвина. Детальный путь образования рибулозо-5- фосфата из трехуглеродных и шестиуглеродных сахаров на этой схеме не показан

19.17. Три АТР и два NADPH доводят СO2 до уровня гексозы
Каков расход энергии для синтеза гексозы? Требуется шесть оборотов цикла Кальвина, поскольку в каждом из них восстанавливается один атом углерода (рис. 19.22). 12 АТР расходуются на фосфорилирование 12 молекул 3-фосфоглицерата в 1,3-бисфосфо- глицерат и 12 NADPH потребляются при восстановлении 12 молекул 1,3-бисфосфо- глицерата в глицеральдегид-3-фосфат. Еще шесть АТР расходуется на регенерирование рибулозо-1,5-бисфосфата.
Теперь напишем уравнение равновесия для суммарной реакции цикла Кальвина:
6СO2 + 18АТР + 12NADPH + 12Н2O → С6Н12O6 + 18ADP + 18Pi + 12NADP+ + 6Н + .
Таким образом, при превращении СO2 в гексозу (глюкозу или фруктозу) потребляются три молекулы АТР и две молекулы NADPH. Эффективность фотосинтеза может быть измерена следующим образом:
1. ∆G 0 для восстановления СO2 до уровня гексозы составляет 114 ккал/моль.
2. При восстановлении NADP + происходит перенос двух электронов. Следовательно, образование двух NADPH требует засасывания фотосистемой 1 четырех фотонов. Электроны, отдаваемые фотосистемой I, восполняются фотосистемой II, которая должна поглотить эквивалентное количество протонов. Значит, для генерирования, требуемого NADPH необходимы восемь фотонов. Одновременно образуются три молекулы АТР, которые нужны для превращения СO2 в гексозу.
3. Содержание энергии в моле фотонов с длиной волны 600 нм равно 47,6 ккал, и, следовательно, приток энергии от восьми молей фотонов составит 381 ккал. Таким образом, общая эффективность фотосинтеза при стандартных условиях составляет минимум 114/381, или 30%.
Биологическая библиотека — материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2023 Все права на дизайн сайта принадлежат С.Є.А.
Источник